
視床下部
最後に見直したもの: 07.07.2025

すべてのiLiveコンテンツは、可能な限り事実上の正確さを保証するために医学的にレビューまたは事実確認されています。
厳格な調達ガイドラインがあり、評判の良いメディアサイト、学術研究機関、そして可能であれば医学的に査読された研究のみにリンクしています。 かっこ内の数字([1]、[2]など)は、これらの研究へのクリック可能なリンクです。
当社のコンテンツのいずれかが不正確、期限切れ、またはその他の疑問があると思われる場合は、それを選択してCtrl + Enterキーを押してください。
視床下部は間脳の下部を形成し、第三脳室底の形成に関与しています。視床下部には、視交叉、視索、漏斗部を伴う灰白質結節、および乳頭体が含まれます。
視交叉(キアズマ・オプティカム)は、視神経(第2頭蓋神経対)の線維によって形成される横方向の隆起であり、部分的に反対側へ交差しています(交差を形成します)。この隆起は、両側で外側および後方に続き、視索(トラトゥス・オプティカス)へと続きます。視索は前穿孔物質から内側および後方に位置し、外側から大脳脚を迂回して屈曲し、皮質下視覚中枢の2つの根で終わります。太い側根(ラディックス・ラテラルリス)は外側膝状体に近づき、細い内側根(ラディックス・メディアリス)は中脳天井の上丘へと続きます。
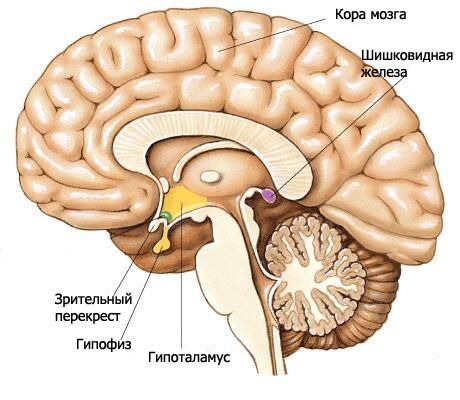
終脳に属する終末板は、視交叉の前面に隣接し、視交叉と癒合している。終末板は大脳縦溝の前部を閉鎖し、薄い灰白質層から構成され、板の側方部では大脳半球の前頭葉実質へと続いている。
視交叉の背後には灰白質結節(tuber cinereum)があり、その背後には乳頭体が、そしてその両側には視索が位置しています。灰白質結節は下方で漏斗(infundibulum)へと進み、漏斗は下垂体へと繋がっています。灰白質結節の壁は、灰白質結節核(nuclei tuberales)を含む薄い灰白質板で形成されています。第三脳室腔の側面からは、漏斗の狭まる窪みが灰白質結節の領域へと突き出し、さらに漏斗へと続いています。
乳頭体(乳頭小体)は、前方の灰白質結節と後方の後孔物質の間に位置しています。乳頭小体は、直径約0.5cmの2つの小さな球状の白色構造物です。白質は乳頭小体の外側のみに位置しています。乳頭小体の内部には灰白質があり、その中で乳頭小体の内側核と外側核(乳頭体核、内側核と外側核)が区別されています。円蓋の柱は乳頭小体で終わっています。
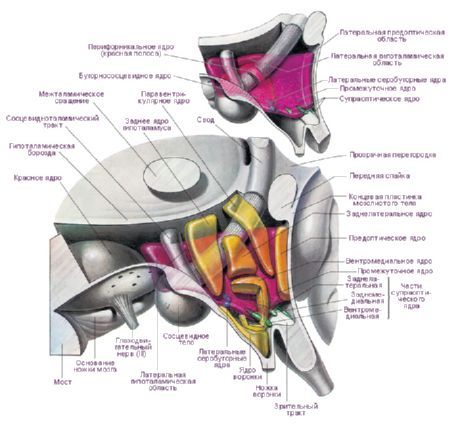
視床下部には、前部(regio hypothalamica anterior)、中間部(regio hypothalamica intermedia)、後部(regio hypothalamica posterior)という3つの主要な視床下部領域があり、それぞれ異なる形状と大きさの神経細胞群が集まって形成されています。これらの領域にある神経細胞群は、30個以上の視床下部核を形成しています。
視床下部核の神経細胞は分泌物(神経分泌物)を産生する能力を有し、この分泌物はこれらの細胞の突起を介して下垂体へ輸送されます。このような核は視床下部神経分泌核と呼ばれます。視床下部の前部には、視索上核(nucleus supraopticus)と室傍核(nuclei paraventriculares)があります。これらの核の細胞の突起は視床下部-下垂体束を形成し、下垂体後葉で終わります。視床下部後部の核群のうち、最も大きいのは、乳頭体内側核と外側核(乳頭体核の内側核と外側核)、および後視床下部核(後視床下部核)です。視床下部中間部の核群には、視床下部下内側核と上内側核(視床下部腹内側核と背内側核)、視床下部背側核(視床下部背側核)、漏斗部核(漏斗部核)、灰白質結節核(結節核)などが含まれます。
視床下部の核は、求心性および遠心性の経路からなる非常に複雑なシステムによって連結されています。そのため、視床下部は身体の多くの栄養機能に調節的な作用を及ぼします。視床下部の核からの神経分泌は、下垂体の腺細胞の機能に影響を与え、いくつかのホルモンの分泌を増加または抑制し、それによって他の内分泌腺の活動を調節します。
視床下部核と下垂体の間に神経と体液のつながりが存在するため、それらを視床下部-下垂体系に結合することが可能になりました。
系統発生学的研究により、視床下部は全ての脊索動物に存在し、両生類ではよく発達しており、爬虫類と魚類ではさらに発達していることが示されています。鳥類は核の分化を明確に示しています。哺乳類では灰白質が高度に発達し、その細胞は核と野に分化します。ヒトの視床下部は、高等哺乳類の視床下部と大きな違いはありません。
視床下部核には多数の分類法があります。E. Gruntelは15対の核を同定し、W. Le Gros Clarkは16対、H. Kuhlenbekは29対の核を同定しました。最も広く用いられている分類法はW. Le Gros Clarkの分類法です。I.N. Bogolepova (1968)は、上記の分類に基づき、個体発生データを考慮して、視床下部核を4つのセクションに分割することを提案しています。
- 前部または吻側部(視索前野と前部群を組み合わせたもの - W. Le Gros Clark) - 視索前内側部および外側部、視交叉上核、視索上核、室傍核、前視床下部野。
- 中部内側部 - 腹内側核、背内側核、漏斗核、後視床下部領域;
- 中部外側部 - 外側視床下部領域、外側視床下部核、結節外側核、結節乳頭核、脳弓周囲核;
- 後部、または乳頭部 - 内側乳頭核、外側乳頭核。
視床下部の解剖学的接続もまた、その(機能的)重要性を明確に示しています。最も重要な求心性経路としては、以下のものが挙げられます。
- 内側前脳束。外側部分は視床下部を嗅球および結節、扁桃周囲領域および海馬に結び付け、内側部分は中隔、対角領域および尾状核に結び付けます。
- 扁桃腺から視床下部の前部まで走る終末帯。
- 海馬から乳頭体まで脳弓を通過する繊維。
- 視床、線条体、淡蒼球視床下部のつながり;
- 脳幹から - 中心被蓋路;
- 大脳皮質(眼窩部、側頭部、頭頂部)から。
したがって、求心性伝達の主な源は、前脳の辺縁系と脳幹の網様体です。
視床下部の流出系も 3 つの方向に分類できます。
- 網様体および脊髄への下行系 - 中脳(縦後束)で終わる神経線維の脳室周囲系(尾部体幹および脊髄の自律神経中枢)、および乳頭体から中脳の網様体に至る乳頭被蓋束。
- 閉鎖機能的大脳辺縁系の一部である乳頭体(乳頭視床束)から視床への経路。
- 下垂体への経路 - 室傍核(繊維の 10~20%)および視索上核(80~90%)から下垂体の後葉および部分的に中葉に至る視床下部-下垂体経路、腹内側核および漏斗核から下垂体前葉に至る結節下垂体経路。
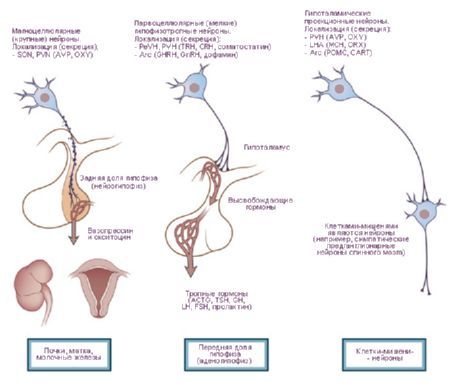
J. ランソン (1935) と W. ヘス (1930, 1954, 1968) の研究では、視床下部刺激による瞳孔の散大と散小、動脈圧の上昇と下降、脈拍の加速と減速に関するデータが提示されました。これらの研究に基づき、交感神経(視床下部後部)と副交感神経(視床下部前部)の作用を及ぼす領域が特定され、視床下部自体が内臓系(臓器や組織に神経支配する)の活動を統合する中枢であると考えられるようになりました。しかし、これらの研究が進むにつれて、特に動物の自由行動において、多くの身体的作用も明らかになりました [Gellhorn E., 1948]。 OG Baklavadzhan(1969)は、視床下部の様々な部位を刺激したところ、大脳皮質の活性化反応、脊髄の単シナプス電位の促進、血圧の上昇が見られるケースもあれば、逆の効果が見られるケースもありました。この場合、栄養反応の閾値が最も高かったのです。O. Sager(1962)は、視床下部のジアテルミー中にY系とEEG同期が抑制され、過度の加熱中に逆の効果が現れることを発見しました。視床下部は、調節機構間の相互作用、体性活動と栄養活動の統合を行う脳の一部であるという考えが定着しました。この観点から、視床下部を交感神経と副交感神経の部分に分けるのではなく、視床下部内のダイナモジェニック(エルゴトロピックとトロフォトロピック)ゾーンを区別する方が正確です。この分類は機能的かつ生物学的な性質を持ち、視床下部が全体的な行動行為の遂行に関与していることを反映しています。言うまでもなく、栄養系だけでなく体性系も恒常性の維持に関与しています。エルゴトロピック領域と栄養向性領域は視床下部のあらゆる部位に存在し、一部の領域では互いに重なり合っています。同時に、それらの「集中」領域を特定することも可能です。例えば、前部(視索前野)では栄養向性装置がより明確に表れ、後部(乳頭体)ではエルゴトロピック装置がより明確に表れています。視床下部と大脳辺縁系および網様体系との主要な求心性および遠心性結合を分析することで、統合的な行動形態の組織化における視床下部の役割が明らかになります。視床下部は、これらの組織の中心に位置するという地理的位置と、生理学的特徴の両方から、このシステムにおいて特別な、つまり中心的な位置を占めています。後者は、脳の特別に構築された部分としての視床下部の役割によって決定され、視床下部は体の内部環境の変化に特に敏感であり、体液指標のわずかな変動にも反応し、これらの変化に応じて適切な行動を形成します。視床下部の特別な役割は、解剖学的および機能的に下垂体に近いことから決定されます。視床下部の核は、特異的核と非特異的核に分けられます。最初のグループには下垂体へ投射する形成物が含まれ、残りは他の核を含み、その刺激の効果は衝撃の強さに応じて変化する可能性があります。視床下部の特異的核は明確な効果を持ち、神経分泌能力において他の脳形成物とは異なります。これには、灰白質結節の視索上核、室傍核、および小細胞核が含まれます。抗利尿ホルモン(ADH)は、視索上核と室傍核で生成され、視床下部-下垂体路の軸索に沿って下垂体後葉まで下降することが確立されています。その後、視床下部のニューロンで放出因子が形成され、それが下垂体前葉に入り、副腎皮質刺激ホルモン(ACTH)、黄体形成ホルモン(LH)、卵胞刺激ホルモン(FSH)、甲状腺刺激ホルモン(TSH)の三大ホルモンの分泌を調節することが示されました。ACTHとTSHの放出因子の形成領域は、正中隆起の前部と視索前野の核であり、GTGの放出因子の形成領域は灰白質結節の後部です。ヒトの視床下部-下垂体束には約100万本の神経線維が含まれていることが確認されています。
脳の他の部位(側頭葉の内側基底構造、脳幹の網様体)も神経内分泌調節に関与していることは疑いようがありません。しかし、最も特異的な器官は視床下部であり、そこには身体の統合反応系、特にストレス性反応に関わる内分泌腺が含まれます。栄養栄養系およびエルゴトロピック系は、活動を確保するために末梢交感神経系と副交感神経系だけでなく、特異的な神経ホルモン装置も利用しています。フィードバック原理に基づいて機能する視床下部-下垂体系は、概ね自己調節的です。実施因子の形成活性は、末梢血中のホルモン濃度によっても決定されます。
このように、視床下部は脳の辺縁系と網様体の重要な構成要素ですが、これらの系に含まれているため、内部環境の変化に対する特別な感受性という形で特定の「入力」を保持し、視床下部-下垂体系、その下にある栄養体への室傍接続、および脳幹の視床と網様体を介して皮質と脊髄に特定の「出力」を保持しています。
 [
[ 何を調べる必要がありますか?
どのように調べる?