
カテコールアミンの合成、分泌および代謝
最後に見直したもの: 19.10.2021

すべてのiLiveコンテンツは、可能な限り事実上の正確さを保証するために医学的にレビューまたは事実確認されています。
厳格な調達ガイドラインがあり、評判の良いメディアサイト、学術研究機関、そして可能であれば医学的に査読された研究のみにリンクしています。 かっこ内の数字([1]、[2]など)は、これらの研究へのクリック可能なリンクです。
当社のコンテンツのいずれかが不正確、期限切れ、またはその他の疑問があると思われる場合は、それを選択してCtrl + Enterキーを押してください。
副腎の大脳層は、構造のステロイドから遠い化合物を産生する。それらは3,4-ジヒドロキシフェニル(カテコール)核を含み、カテコールアミンと呼ばれる。これらには、アドレナリン、ノルエピネフリンおよびドーパミンβ-オキシチラミンが含まれる。
シーケンスカテコールアミン合成は非常に簡単です:チロシン→のジヒドロキシ(DOPA)→→ドーパミン、ノルエピネフリン→アドレナリン。チロシンは食物と共に体内に入るが、フェニルアラニンヒドロキシラーゼの作用下で肝臓中のフェニルアラニンからも形成することができる。組織におけるチロシンの最終生成物は異なる。中枢神経系カテコールアミン、ドーパミン合成完成形態の特定のニューロンにおいて、ノルアドレナリン - 副腎髄質の処理は、交感神経の端部に、アドレナリンの形成に進みます。
チロシンのDOPAへの変換はチロシンヒドロキシラーゼによって触媒され、補因子はテトラヒドロ - ビオプテリンおよび酸素である。カテコールアミン生合成の全プロセスの速度を制限し、このプロセスの最終生成物によって阻害されるのはこの酵素であると考えられている。チロシンヒドロキシラーゼは、カテコールアミンの生合成に対する調節効果の主要な目的である。
比較的非特異的及び脱炭酸され、そして他の芳香族L-アミノ酸 - ドーパミンへの変換ドーパは、酵素ドーパ - デカルボキシラーゼ(ピリドキサール補因子)によって触媒されます。しかしながら、活性およびこの酵素を変化させることによってカテコールアミンの合成を修飾する可能性が示唆されている。いくつかのニューロンでは、ドーパミンのさらなる変換のための酵素は存在せず、最終生成物である。他の組織には、ドーパミンをノルアドレナリンに変換するドーパミンベータヒドロキシラーゼ(補因子は銅、アスコルビン酸および酸素)が含まれています。ノルアドレナリンからアドレナリン形成メチル - 副腎髄質(ただし、交感神経終末)に存在フェニルエタノールアミンです。この場合のメチル基の供与体はS-アデノシルメチオニンである。
フェニルエタノールアミン-N-Metiltransferazyの合成は、ポータル静脈系の大脳皮質の層に落ちるグルココルチコイドによって誘導されたことを覚えておくことが重要です。これは、1人の体に二つの異なる内分泌腺を組み合わせた事実嘘を説明することがあります。血液細胞は、静脈洞から本質的adrenalinprodutsiruyuschieを得副腎皮質に局在している間に、動脈血管の周囲に配置され、ノルエピネフリンを製造副腎髄質細胞という事実によって強調アドレナリンのグルココルチコイド合成の意味。
カテコール-O-メチルトランスフェラーゼ(COMT)およびモノアミンオキシダーゼ(MAO):カテコールアミンの崩壊は、主に2つの酵素システムの影響下で起こります。図1に概略的に示される減衰のエピネフリン及びノルエピネフリンの主な方法。54メチル基のS-adrenozilmetioninaカテコールアミンの供与体の存在下でCOMTの作用下でMAOの影響下でバニリルマンデルに(アルデヒドの存在下で)アルデヒドよりに変換ノルメタネフリンのメタネフリン(エピネフリンおよびノルエピネフリンの3-O-メチル誘導体)に変換酸(ICH) - ノルアドレナリンおよびアドレナリンの主分解物。3,4- dioksimindalnuyu酸とIUD - 最初のカテコールアミンのMAOの作用にさらされたときに同じ場合に、COMT、それらはアルデヒド及びCOMTの影響を受け、次いで3,4- dioksimindalevyアルデヒドに変換されません。カテコールアミンのアルコールデヒドロゲナーゼの存在下で、3-メトキシ-4- oksifenilglikol、CNSにおけるエピネフリンおよびノルエピネフリンの劣化の主な最終製品を形成することができます。
崩壊ドーパミンは、その代謝物は、β-炭素原子上にヒドロキシル基を欠いていることを除いて、そのため、代わりにバニリル - マンデル酸形成ホモバニリン(HVA)及び3-メトキシ-4- oksifeniluksusnaya酸、同様に進行します。
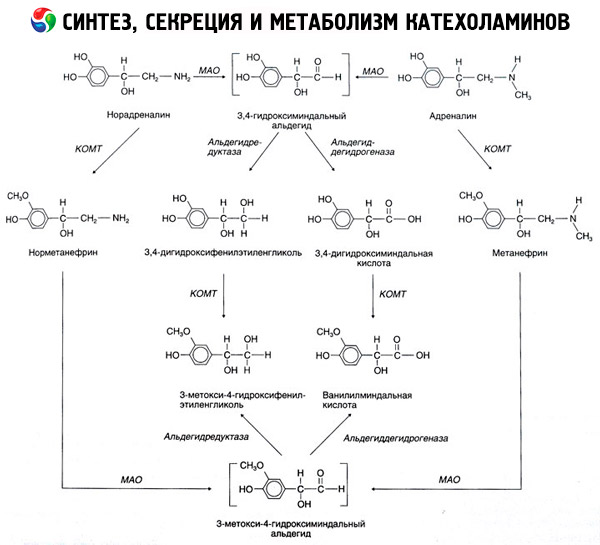
顕著な生物学的活性を有する中間体生成物であるカテコールアミンの分子の酸化のためのキノイド経路の存在も仮定することができる。
交感神経末端、副腎髄質における細胞質ゾル酵素、アドレナリンやノルアドレナリンの作用によって形成され、分解酵素の作用から保護する分泌顆粒を入力してください。カテコールアミンを顆粒で捕捉するには、エネルギーコストが必要です。細胞質内顆粒からのホルモンの拡散を防止クロモグラニン - 及び特定のタンパク質:(1〜4の比で)しっかりとATPに結合副腎髄質カテコールアミンのクロマフィン顆粒中。
(細胞外液中に - - カテコールアミン、ドーパミンベータヒドロキシラーゼ、ATPおよびクロモグラニン融合膜細胞表面と顆粒および可溶分の全体的な収率とのギャップ)カテコールアミンの分泌への直接刺激は明らかにカルシウムの細胞は、エキソサイトーシスを刺激浸透さ。
カテコールアミンの生理作用とその作用機序
カテコールアミンの作用は、標的細胞の特異的受容体との相互作用から始まる。甲状腺およびステロイドホルモンの受容体は細胞内に局在化している場合は、カテコールアミン受容体(ならびにアセチルコリンおよびペプチドホルモン)は、外側細胞表面上に存在します。
長い間、他のエフェクトは、イソプロテレノール、エピネフリンまたはノルエピネフリンの作用に優れている間、いくつかの反応アドレナリンやノルアドレナリンの点で、合成カテコールアミンイソプロテレノールよりも有効であることが確立されています。これに基づいて、アルファおよびベータの2種類のアドレナリン受容体の組織における存在に関する概念が開発され、その中には、これらの2つの型のうちの1つのみが存在し得るものもある。イソプロテレノールは、合成化合物フェニレフリン一方、β-アドレナリン受容体の最も強力なアゴニストである - アルファ - アドレナリン受容体の最も強力なアゴニスト。ナチュラルカテコールアミン - アドレナリンとノルアドレナリンは - 両方のタイプの受容体と相互作用することができるが、アドレナリンはベータ版のための高い親和性を取り、ノルエピネフリン - アルファ受容体。
ベータ1受容体(心臓、脂肪細胞)およびベータ2受容体(気管支、血管、など...):カテコールアミンが強くサブタイプに分け、ベータ型が可能、平滑筋のβ受容体よりも心臓のβアドレナリン受容体を活性化します。アドレナリンおよびノルアドレナリンのベータ1受容体の優れた作用にイソプロテレノールの作用のみが10倍、それは天然のカテコールアミンよりも100~1000倍強力に作用するベータ2受容体があります。
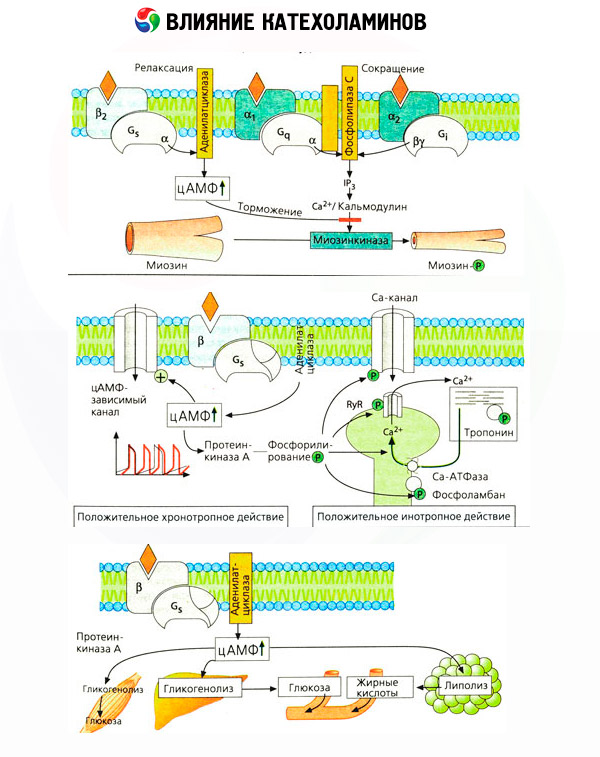
特定のアンタゴニスト(ベータ受容体に対してアルファおよびプロプラノロールに対するフェントラミンおよびフェノキシベンザミン)の使用は、アドレナリン受容体の分類の妥当性を確認した。ドーパミンは、α-およびβ-受容体の両方と相互作用することができるが、様々な組織(脳、下垂体、船舶)にハロペリドールある特定のブロッカーを発見し、自分のドーパミン作動性受容体。ベータ受容体の数は、細胞あたり1000〜2000の範囲である。ベータ受容体によって媒介されるカテコールアミンの生物学的効果は、原則として、アデニレートシクラーゼの活性化およびcAMPの細胞内含量の増加に関連する。受容体と酵素は機能的には結合しているが、異なる巨大分子を表している。アデニル酸シクラーゼ活性の調節において、ホルモン受容体複合体の影響下で、グアノシン三リン酸(GTP)および他のプリンヌクレオチドが関与する。酵素の活性を増加させることにより、それらはアゴニストに対するベータ受容体の親和性を低下させるようである。
【0003】除神経構造の感度を増加させる現象は、長い間知られている。逆に、アゴニストへの長期間の暴露は、標的組織の感受性を低下させる。ベータ受容体の研究は、これらの現象を説明することを可能にした。イソプロテレノールの長期作用は、ベータ受容体の数の減少によるアデニレートシクラーゼの感受性の喪失につながることが示された。
脱感作のプロセスは、タンパク質合成の活性化を必要とせず、恐らく不可逆的なホルモン - 受容体複合体の漸進的形成によるものである。対照的に、交感神経末端を破壊する6-オキシドファaminの投与は、組織におけるβ受容体の反応の数の増加を伴う。交感神経活動の増加が、カテコールアミンに関連した血管および脂肪組織の加齢関連脱感作を決定することは排除されない。
異なる器官のアドレナリン受容体の数は、他のホルモンによって制御することができる。したがって、それはエストラジオール、プロゲステロンおよびカテコールアミンに対する収縮応答の対応する増減を伴う子宮内アルファ - アドレナリン受容体の数を減らすことを増加させます。細胞内の「セカンドメッセンジャー」は、ベータ受容体アゴニストの作用によって形成された場合は、確実にcAMPは、α-アドレナリン作動性効果の送信に関連して、あることはより複雑です。様々な機序があると考えられている:cAMPのレベルの低下、cAMPの含有量の増加、カルシウムの細胞動態の調節、
体内でさまざまな効果を再現するためには、通常、ノルエピネフリンより5〜10倍低いエピネフリンの用量が必要です。後者はA-およびベータ - アドレナリン受容体に対してより効果的であるが、α-およびβ-受容体の両方と相互作用することができる内因性カテコールアミンの両方ということを覚えておくことが重要です。したがって、この身体のアドレナリン作動性への生物学的応答は、その中に存在するレセプターのタイプに大きく依存する。しかしながら、これは、交感神経副腎系の神経または体液結合の選択的活性化が不可能であることを意味するものではない。ほとんどの場合、さまざまなリンクの活動が強化されています。従って、血圧(起立性低血圧)の低下は、交感神経終末からのノルエピネフリン放出を主に伴う一方、それは、反射低血糖副腎髄質を活性化することが想定されます。
アドレナリン受容体および種々の組織におけるそれらの活性化の効果
|
システム、オルガン |
アドレナリン受容体型 |
反応 |
|
心臓血管系: |
||
|
心臓 |
ベータ |
収縮、伝導および収縮の頻度の増加 |
|
細動脈: |
||
|
皮膚および粘膜 |
アルファ |
削減 |
|
骨格筋 |
ベータ |
拡張機能の削減 |
|
腹部の器官 |
アルファ(もっと) |
削減 |
|
ベータ |
拡張 |
|
|
静脈 |
アルファ |
削減 |
|
呼吸器系: |
||
|
気管支筋 |
ベータ |
拡張 |
|
消化器系: |
||
|
胃 |
ベータ |
モーター機能の低下 |
|
腸 |
アルファ |
括約筋の減少 |
|
脾臓 |
アルファ |
削減 |
|
ベータ |
リラクゼーション |
|
|
膵臓の外部秘密の部分 |
アルファ |
分泌の減少 |
|
泌尿生殖器系: |
アルファ |
括約筋の縮小 |
|
膀胱 |
ベータ |
エクソシストの筋肉をリラックス |
|
男性の性的臓器 |
アルファ |
射精 |
|
目 |
アルファ |
瞳孔が拡張 |
|
レザー |
アルファ |
発汗の増加 |
|
唾液腺 |
アルファ |
カリウムと水の分離 |
|
ベータ |
アミラーゼの分泌 |
|
|
内分泌腺: |
||
|
膵島 |
||
|
ベータ細胞 |
アルファ(もっと) |
インスリン分泌の減少 |
|
ベータ |
インスリン分泌の増加 |
|
|
アルファ細胞 |
ベータ |
グルカゴンの分泌増加 |
|
8細胞 |
ベータ |
ソマトスタチンの分泌増加 |
|
視床下部や下垂体: |
||
|
体動虫 |
アルファ |
STH分泌の増加 |
|
ベータ |
STH分泌の減少 |
|
|
ラクトトロファ |
アルファ |
プロラクチン分泌の減少 |
|
甲状腺刺激ホルモン |
アルファ |
TSH分泌の減少 |
|
コルチコトロフス |
アルファ |
ACTHの分泌増加 |
| ベータ | ACTHの分泌低下 | |
|
甲状腺: |
||
|
濾胞細胞 |
アルファ |
サイロキシンの分泌低下 |
|
ベータ |
サイロキシン分泌の増加 |
|
|
傍胞子(K)細胞 |
ベータ |
カルシトニンの分泌増加 |
|
副甲状腺 |
ベータ |
PTHの分泌増加 |
|
腎臓 |
ベータ |
レニン分泌の増加 |
|
胃 |
ベータ |
ガストリン分泌を増加させる |
|
基本交換 |
ベータ |
酸素消費量の増加 |
|
肝臓 |
? |
グルコース収率からのグリコーゲン分解および糖新生の増加; ケトン体の放出によるケトン生成の増加 |
|
脂肪組織 |
ベータ |
遊離脂肪酸およびグリセロールの放出による脂肪分解の増加 |
|
骨格筋 |
ベータ |
ピルビン酸塩および乳酸塩の放出による解糖の増加; アラニン、グルタミンの収率の低下を伴うタンパク質分解の減少 |
カテコールアミンの静脈内投与の結果が常に適切に内因性化合物の影響を反映していないことに留意することが重要です。それは主に体内ない血液中に立っているが、直接シナプス間隙にこれは、主にノルアドレナリンに適用されます。従って、内因性ノルエピネフリン活性化、例えば、のみならず(迷走神経を介して)血管アルファ受容体ノルアドレナリン外部リードの投与に対し、主血管アルファ受容体と反射の活性化(血圧上昇)も、心臓ベータ受容体(動悸)、減速動悸。
低用量のエピネフリンは、主に筋肉血管および心臓のβ受容体を活性化し、末梢血管抵抗の低下をもたらし、心臓の微小体積を増加させる。場合によっては、第1の効果が優勢であり、アドレナリンの投与後に低血圧が発現する。高用量では、アドレナリンはまた、末梢血管抵抗の増加を伴うアルファ受容体を活性化し、心臓の微小体積の増加の背景に対して、血圧の上昇をもたらす。しかしながら、血管ベータ受容体に対するその効果もまた保存されている。その結果、収縮期圧の上昇は、拡張期圧の類似の値(脈圧の上昇)を超える。さらに多くの用量が導入されると、エピネフリンのアルファミメティック効果、すなわち、ノルアドレナリンの影響下での収縮期および拡張期の圧力上昇が並行して増加し始める。
カテコールアミンが代謝に与える影響は、その直接的および間接的な影響から成り立っています。第1のものは、主にβ受容体を介して実現される。より複雑なプロセスが肝臓と関連している。肝グリコーゲン分解の増強は伝統的にβ受容体活性化の結果と考えられてきたが、これにはα受容体の関与に関するデータもある。カテコールアミンの仲介された作用は、多くの他のホルモン、例えばインスリンの分泌の調節に関連する。その分泌に対するアドレナリンの作用において、アルファ - アドレナリン作動性成分は明らかに優勢であり、何らかのストレスがインスリン分泌の阻害を伴うことが示されているからである。
カテコールアミンの直接作用および間接作用の組合せは、グルコースの肝臓産生の増加だけでなく、末梢組織によるその利用の阻害とも関連する高血糖を引き起こす。脂肪分解の促進は、脂肪酸への肝臓への送達の増加およびケトン体の生成の強化を伴う高脂血症を引き起こす。筋肉の解糖の強化は、一緒に脂肪組織から遊離グリセロールと血中乳酸およびピルビン酸で出力増大につながる、肝臓の糖新生の前任者です。
カテコールアミンの分泌の調節。製品および交感神経系および副腎髄質の応答の方法の類似性は、単体交感神経副腎系のリリース神経及びホルモンのリンクにこれらの構造を結合するための基礎でした。II-IIIの腰椎セグメント - 種々の求心性信号は、子宮頸部VIIIのレベルで脊髄の横方向の角に位置する節前ニューロンの細胞体上の遠心性パーセルスイッチングを発し、そこから脊髄及び延髄の視床下部及び中心に集中しています。
これらの細胞の節前軸索は、交感神経鎖、又は副腎髄質細胞の神経節に局在している脊髄を出てニューロンとシナプス結合を形成しています。これらは、コリン作動性節前繊維です。節後交感神経ニューロンおよび副腎髄質クロム親和性細胞の第一の基本的な違いは、後者が血液にアドレナリン作動性化合物を強調することによって神経伝導(節後アドレナリン作動性神経)および体液性がコリン作動性入力信号に伝達されることにあります。第2の相違点は、ノルエピネフリンを生成節後神経に低減される副腎髄質細胞つつ - 好ましくアドレナリン。これらの2つの物質は、組織に対して異なる効果を有する。