
下垂体
最後に見直したもの: 04.07.2025

すべてのiLiveコンテンツは、可能な限り事実上の正確さを保証するために医学的にレビューまたは事実確認されています。
厳格な調達ガイドラインがあり、評判の良いメディアサイト、学術研究機関、そして可能であれば医学的に査読された研究のみにリンクしています。 かっこ内の数字([1]、[2]など)は、これらの研究へのクリック可能なリンクです。
当社のコンテンツのいずれかが不正確、期限切れ、またはその他の疑問があると思われる場合は、それを選択してCtrl + Enterキーを押してください。
下垂体(下垂体、s.glandula pituitaria)は、蝶形骨のトルコ鞍の下垂体窩に位置し、脳の硬膜突起によって頭蓋腔から分離され、トルコ鞍横隔膜を形成します。この横隔膜の開口部を通して、下垂体は間脳視床下部の漏斗部に接続されます。下垂体の横幅は10~17 mm、前後幅は5~15 mm、縦幅は5~10 mmです。下垂体の質量は、男性で約0.5 g、女性で約0.6 gです。下垂体は外側から被膜で覆われています。
下垂体は2つの異なる原基から発達するため、器官には前葉と後葉の2つの葉が区別されます。下垂体前葉(adenohypophysis、s.lobus anterior)はより大きく、下垂体全体の質量の70~80%を占めます。前葉は後葉よりも密度が高いです。前葉には、下垂体窩の前部を占める遠位部(pars trailis)、後葉との境界に位置する中間部(pars intermedia)、および上方に伸びて視床下部の漏斗に接続する結節部(pars tuberalis)が区別されます。血管が豊富なため、前葉は赤みがかった淡黄色をしています。下垂体前葉の実質は、数種類の腺細胞によって構成され、それらの細胞間には類洞毛細血管が分布しています。下垂体前葉細胞の半分(50%)は好色性腺細胞で、細胞質内にクロム塩でよく染まる細粒を有しています。これらは好酸性腺細胞(下垂体前葉細胞の40%)と好塩基性腺細胞(10%)です。好塩基性腺細胞には、性腺刺激ホルモン産生細胞、副腎皮質刺激ホルモン産生細胞、甲状腺刺激ホルモン産生細胞が含まれます。好色性腺細胞は小型で、核が大きく細胞質は少量です。これらの細胞は好色性腺細胞の前駆細胞であると考えられています。下垂体前葉細胞の残りの50%は好色性腺細胞です。
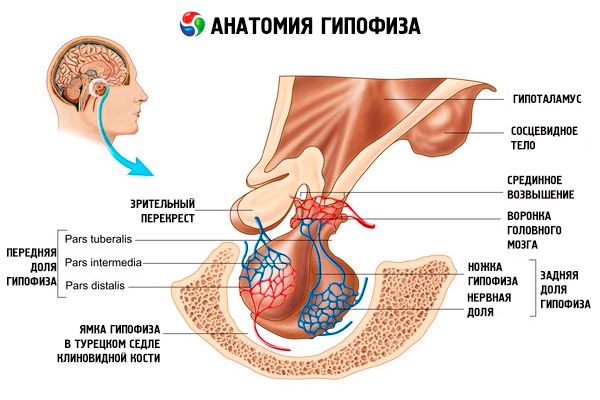
神経下垂体後葉(neurohypophysis、s.lobus posterior)は、下垂体窩の後部に位置する神経葉(lobus nervosus)と、下垂体前葉の結節部の後ろに位置する漏斗部(infundibulum)から構成されます。下垂体後葉は、神経グリア細胞(下垂体細胞)、視床下部の神経分泌核から神経下垂体前葉へと伸びる神経線維、そして神経分泌小体によって形成されます。
下垂体は、神経線維(経路)と血管を介して、間脳の視床下部と機能的に結合しており、視床下部は下垂体の活動を調節します。下垂体と視床下部は、神経内分泌、血管、神経系の接続とともに、通常、視床下部-下垂体系と考えられています。
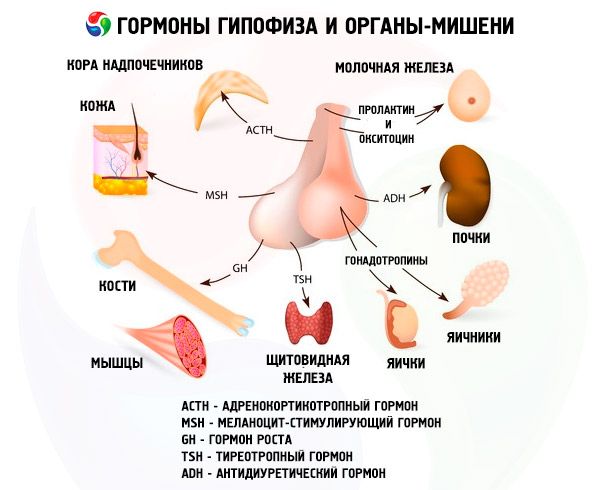
下垂体前葉および下垂体後葉のホルモンは、主に他の内分泌腺を介して、体の多くの機能に影響を与えます。下垂体前葉では、好酸性腺細胞(アルファ細胞)が成長ホルモン(ソマトトロピンホルモン)を産生し、幼若生物の成長と発達過程の調節に関与します。副腎皮質刺激ホルモン(ACTH)を分泌する副腎皮質刺激ホルモン(ACTH)は、副腎からのステロイドホルモンの分泌を刺激します。甲状腺刺激ホルモン(TSH)を分泌する甲状腺刺激ホルモン(TSH)は、甲状腺の発達に影響を与え、甲状腺ホルモンの産生を活性化します。性腺刺激ホルモン(FSH)、黄体形成ホルモン(LH)、プロラクチンは、身体の性成熟に影響を与え、卵巣における卵胞の発育、排卵、乳腺の成長と女性における乳汁分泌、男性における精子形成過程を調節・刺激します。これらのホルモンは、好塩基性腺細胞(β細胞)によって産生されます。下垂体脂肪親和性因子もこの部位から分泌され、体内の脂肪の動員と利用に影響します。前葉の中間部では、メラノサイト刺激ホルモンが生成され、体内の色素(メラニン)の形成を制御します。
視床下部の視索上核と室傍核の神経分泌細胞は、バソプレシンとオキシトシンを産生します。これらのホルモンは、視床下部-下垂体路を構成する軸索に沿って下垂体後葉の細胞へと輸送されます。下垂体後葉からこれらの物質は血液中に入ります。バソプレシンというホルモンには血管収縮作用と抗利尿作用があり、抗利尿ホルモン(ADH)とも呼ばれます。オキシトシンは子宮筋の収縮を刺激し、授乳中の乳腺からの乳汁分泌を促進し、黄体の発達と機能を抑制し、消化管の平滑筋(非横紋筋)の緊張変化に影響を与えます。
下垂体の発達
下垂体前葉は、口側湾背壁上皮から環状の突起(ラトケ嚢)として発達する。この外胚葉の突起は、将来の第三脳室の底部に向かって成長する。その突起に向かって、第二大脳胞(将来の第三脳室の底部)の下面から突起が成長し、そこから漏斗部の灰白質結節と下垂体後葉が発達する。
下垂体の血管と神経
内頸動脈と大脳動脈輪の血管からは、上下垂体動脈と下下垂体動脈が下垂体へと伸びています。上下垂体動脈は視床下部の灰白質核と漏斗部へと向かい、そこで互いに吻合して脳組織を貫通する毛細血管、すなわち一次毛細血管網を形成します。この網の長短のループから門脈が形成され、下垂体前葉へと伸びています。下垂体前葉の実質では、これらの静脈は幅広い洞毛細血管へと分岐し、二次毛細血管網を形成します。下垂体後葉への血液供給は、主に下垂体下垂体動脈によって行われています。上下垂体動脈と下下垂体動脈の間には長い動脈吻合部があります。二次毛細血管網からの静脈血の流出は、脳硬膜の海綿静脈洞と海綿静脈洞間洞に流入する静脈系を通して行われます。
動脈とともに器官内に入る交感神経線維は、下垂体の神経支配に関与する。節後交感神経線維は内頸動脈神経叢から分岐する。さらに、視床下部核に位置する神経分泌細胞の突起の終末部が下垂体後葉に多数存在する。
下垂体の加齢に伴う特徴
新生児における下垂体の平均重量は0.12gです。この臓器の重量は10歳までに2倍、15歳までに3倍になります。20歳までに下垂体の重量は最大(530~560mg)に達し、その後はほぼ一定です。60歳を過ぎると、この内分泌腺の重量はわずかに減少します。
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
下垂体ホルモン
体内の神経系とホルモン系の調節の統一性は、下垂体と視床下部の密接な解剖学的・機能的連結によって確保されています。この複合体は、内分泌系全体の状態と機能を決定します。
末梢腺の機能を直接制御する多数のペプチドホルモンを産生する主要な内分泌腺は下垂体です。これは赤灰色の豆の形をした構造で、重さ0.5~0.6 gの繊維質の被膜で覆われています。性別や年齢によって若干異なります。下垂体は、発達、構造、機能が異なる2つの葉に分けられるという一般的な考え方は変わりません。前者は遠位部、後部は神経下垂体前葉です。前者は腺の全体積の約70%を占め、従来、遠位部、漏斗部、中間部に分けられ、後者は後部、または葉と下垂体茎に分けられます。下垂体は蝶形骨のトルコ鞍の下垂体窩に位置し、茎を介して脳につながっています。前葉の上部は視交叉と視索で覆われています。下垂体への血液供給は非常に豊富で、内頸動脈の枝(上下垂体動脈と下下垂体動脈)と大脳動脈輪の枝によって行われています。上下垂体動脈は下垂体前葉への血液供給に関与し、下垂体動脈は神経下垂体前葉への血液供給に関与し、視床下部の大細胞核の軸索の神経分泌終末と接触します。前者は視床下部の正中隆起に入り、そこで毛細血管網(一次毛細血管叢)へと分散します。これらの毛細血管(視床下部中基底核の小さな神経分泌細胞の軸索末端が接触する)は門脈に集まり、下垂体柄に沿って下垂体前葉実質へと下降し、そこで再び洞毛細血管網(二次毛細血管叢)へと分岐する。こうして、視床下部中隆起を通過し、視床下部腺下垂体刺激ホルモン(放出ホルモン)が豊富に含まれた血液は、下垂体前葉へと流入する。
下垂体前葉ホルモンを豊富に含んだ血液は、二次神経叢の多数の毛細血管から静脈系を経て流出し、硬膜の静脈洞を経て全身の血流へと流れ込む。このように、視床下部から下行方向に血流が流れる下垂体門脈系は、下垂体前葉の向性機能に対する神経体液性制御という複雑なメカニズムの形態機能的構成要素である。
下垂体は、下垂体動脈に沿う交感神経線維によって支配されています。交感神経線維は、内頸動脈神経叢を通過し、上頸神経節につながる節後線維から生じます。下垂体後葉は視床下部から直接神経支配を受けません。後葉は視床下部の神経分泌核から神経線維を受け取ります。
下垂体前葉は、その組織学的構造が非常に複雑です。好色性と好色性の2種類の腺細胞を有します。後者はさらに好酸性と好塩基性に分類されます(下垂体の詳細な組織学的説明は、本書の該当セクションに記載されています)。しかし、下垂体前葉の実質を構成する腺細胞が産生するホルモンは、その多様性ゆえに、化学的性質がある程度異なり、分泌細胞の微細構造はそれぞれのホルモンの生合成特性に対応している必要があることに留意する必要があります。しかし、下垂体前葉には、複数のホルモンを産生できる腺細胞の移行形態が観察されることがあります。下垂体前葉の腺細胞の種類は、必ずしも遺伝的に決定されているわけではないという証拠があります。
トルコ鞍の横隔膜の下には、前葉の漏斗状の部分があります。この部分は下垂体柄を包み込み、灰白質結節に接しています。下垂体前葉のこの部分は、上皮細胞の存在と豊富な血液供給を特徴としています。また、ホルモン活性も有しています。
下垂体の中間(中央)部分は、数層の大きな分泌活性好塩基性細胞で構成されています。
下垂体はホルモンを介して様々な機能を果たします。前葉では副腎皮質刺激ホルモン(ACTH)、甲状腺刺激ホルモン(TSH)、卵胞刺激ホルモン(FSH)、黄体形成ホルモン(LH)、脂肪親和性ホルモン、そして成長ホルモン(STO)、プロラクチンが産生されます。中葉ではメラノサイト刺激ホルモン(MSH)が、後葉ではバソプレシンとオキシトシンが蓄積されます。
ACTH
下垂体ホルモンは、タンパク質、ペプチドホルモン、および糖タンパク質からなるグループです。下垂体前葉ホルモンの中で、ACTHは最もよく研究されています。ACTHは好塩基球細胞によって産生されます。その主な生理機能は、副腎皮質におけるステロイドホルモンの生合成と分泌を促進することです。ACTHはまた、メラノサイト刺激作用と脂肪親和作用も示します。1953年に純粋に単離されました。その後、ヒトおよび多くの哺乳類において39個のアミノ酸残基からなる化学構造が確立されました。ACTHには種特異性はありません。現在では、ホルモン自体と、天然ホルモンよりも活性の高い様々な分子断片の化学合成が行われています。このホルモンの構造は、2つのペプチド鎖セクションから構成されており、1つはACTHの検出と受容体への結合を確実にし、もう1つは生物学的効果をもたらします。ACTHは、ホルモンと受容体の電荷の相互作用を介してACTH受容体に結合すると考えられています。 ACTH の生物学的エフェクターの役割は、4-10 分子 (Met-Glu-His-Phen-Arg-Tri-Tri) のフラグメントによって実行されます。
ACTHのメラノサイト刺激活性は、分子内に13個のアミノ酸残基からなり、α-メラノサイト刺激ホルモンの構造を繰り返すN末端領域が存在することに起因します。この領域には、他の下垂体ホルモンにも存在するヘプタペプチドが含まれており、副腎皮質刺激作用、メラノサイト刺激作用、および脂肪親和作用を有しています。
ACTHの作用における鍵となる瞬間は、cAMPの関与による細胞質内の酵素タンパク質キナーゼの活性化であると考えられています。リン酸化されたタンパク質キナーゼは酵素エステラーゼを活性化し、コレステロールエステルを脂肪滴中の遊離物質に変換します。リボソームのリン酸化の結果として細胞質で合成されたタンパク質は、遊離コレステロールとシトクロムP-450の結合を刺激し、脂肪滴からミトコンドリアへと輸送します。ミトコンドリアには、コレステロールをコルチコステロイドに変換する酵素がすべて存在します。
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
甲状腺刺激ホルモン
TSH(甲状腺刺激ホルモン)は、甲状腺の発達と機能、そして甲状腺ホルモンの合成と分泌のプロセスの主要な調節因子です。この複合タンパク質(糖タンパク質)は、αサブユニットとβサブユニットで構成されています。最初のサブユニットの構造は、黄体形成ホルモンのαサブユニットと一致しています。さらに、それは異なる動物種でほぼ一致しています。ヒトTSHのβサブユニットのアミノ酸残基配列は解読されており、119個のアミノ酸残基で構成されています。ヒトとウシTSHのβサブユニットは多くの点で類似していることが注目されます。糖タンパク質ホルモンの生物学的特性と生物学的活性の性質は、βサブユニットによって決定されます。また、βサブユニットは、ホルモンが様々な標的臓器の受容体と相互作用することを確実にします。しかし、ほとんどの動物では、βサブユニットは、ホルモンの一種の活性化因子として機能するαサブユニットと結合して初めて特異的な活性を示します。後者は、βサブユニットの特性によって決定される黄体形成、卵胞刺激、および甲状腺刺激作用を等確率で誘導します。発見された類似性から、これらのホルモンは共通の祖先から進化の過程で生じたと結論付けることができます。また、βサブユニットはこれらのホルモンの免疫学的特性も決定します。αサブユニットはβサブユニットをタンパク質分解酵素の作用から保護し、下垂体から末梢標的臓器への輸送を促進すると考えられています。
性腺刺激ホルモン
ゴナドトロピンは、LHとFSHという形で体内に存在します。これらのホルモンの機能的役割は、一般的に男女ともに生殖プロセスを確実に行うことです。TSHと同様に、FSHは複合タンパク質(糖タンパク質)です。FSHは、女性の卵巣で卵胞の成熟を促し、男性の精子形成を刺激します。LHは、女性の卵胞破裂と黄体形成を促し、エストロゲンとプロゲステロンの分泌を刺激します。男性では、同じホルモンが間質組織の発達とアンドロゲンの分泌を促進します。ゴナドトロピンの作用は相互に依存しており、同期して発現します。
女性におけるゴナドトロピン分泌の動態は月経周期中に変化し、十分に詳細に研究されている。排卵前期(卵胞期)には、LH含有量は比較的低く、FSH含有量は増加する。卵胞が成熟するにつれて、エストラジオール分泌が増加し、これが下垂体によるゴナドトロピン産生の増加と、LHとFSHの両方の周期の発生を促す。つまり、性ステロイドはゴナドトロピンの分泌を刺激するのである。
現在、LHの構造は解明されています。TSHと同様に、LHはサブユニットaとbの2つのサブユニットで構成されています。LHのαサブユニットの構造は、様々な動物種においてほぼ一致しており、TSHのαサブユニットの構造と一致しています。
LHのβサブユニットの構造は、TSHのβサブユニットの構造とは大きく異なりますが、4~5個のアミノ酸残基からなるペプチド鎖の4つの同一セクションを有しています。TSHでは、これらのセクションは27~31、51~54、65~68、および78~83番目の位置に局在しています。LHとTSHのβサブユニットはホルモンの特異的な生物学的活性を決定するため、LHとTSHの構造における相同セクションはβサブユニットとαサブユニットの結合を確実にし、構造が異なるセクションはホルモンの生物学的活性の特異性を担っていると考えられます。
天然LHはタンパク質分解酵素の作用に対して非常に安定していますが、βサブユニットはキモトリプシンによってすぐに切断され、αサブユニットは酵素によって加水分解されにくいため、キモトリプシンがペプチド結合にアクセスするのを防ぐ保護的な役割を果たします。
FSHの化学構造については、研究者はまだ決定的な結論を得ていません。LHと同様に、FSHは2つのサブユニットで構成されていますが、FSHのβサブユニットはLHのβサブユニットとは異なります。
プロラクチン
もう一つのホルモンであるプロラクチン(乳汁分泌ホルモン)は、生殖過程において積極的な役割を果たしています。哺乳類におけるプロラクチンの主な生理学的特性は、乳腺の発達と授乳、皮脂腺および内臓の成長を促進するという形で現れます。また、男性におけるステロイドの二次性徴への作用の発現を促進し、マウスやラットにおける黄体の分泌活動を刺激し、脂肪代謝の調節にも関与しています。近年、プロラクチンは母性行動の調節因子として大きな注目を集めており、このような多機能性は進化の過程によって説明されます。プロラクチンは古代の下垂体ホルモンの一つであり、両生類にも存在します。現在、一部の哺乳類種におけるプロラクチンの構造は完全に解読されています。しかし、最近まで、科学者たちはヒトにそのようなホルモンが存在することに疑問を抱いていました。多くの人は、その機能は成長ホルモンによって担われていると信じていました。現在、ヒトにプロラクチンが存在するという説得力のある証拠が得られ、その構造も部分的に解明されています。プロラクチン受容体は成長ホルモンと胎盤性ラクトゲンに活発に結合し、これら3つのホルモンの作用機序は単一であることを示唆しています。
ソマトトロピン
成長ホルモンであるソマトトロピンは、プロラクチンよりもさらに広い作用スペクトルを有します。プロラクチンと同様に、下垂体前葉の好酸性細胞によって産生されます。ソマトトロピンは骨格の成長を刺激し、タンパク質の生合成を活性化し、脂肪の動員作用を有し、体格の増加を促進します。さらに、代謝プロセスを調整します。
後者におけるホルモンの関与は、例えば血糖値が下がったときに下垂体からのホルモン分泌が急増するという事実によって確認されます。
このヒトホルモンの化学構造は、191個のアミノ酸残基から完全に解明されています。その一次構造は、絨毛性ソマトマンモトロピンまたは胎盤性ラクトゲンのものと類似しています。これらのデータは、両ホルモンが生物学的活性に違いはあるものの、進化の過程で著しく近縁であったことを示しています。
問題のホルモンの高い種特異性を強調する必要があります。たとえば、動物由来のSTHはヒトでは不活性です。これは、ヒトと動物のSTH受容体間の反応と、ホルモン自体の構造の両方によって説明されます。現在、STHの複雑な構造の中で生物学的活性を示す活性中心を特定するための研究が進行中です。他の特性を示す分子の個々の断片が研究されています。たとえば、ヒトSTHをペプシンで加水分解した後、14個のアミノ酸残基で構成され、分子セクション31〜44に対応するペプチドが単離されました。これは成長効果はありませんでしたが、脂肪親和性活性において天然ホルモンを大幅に上回りました。ヒト成長ホルモンは、動物の同様のホルモンとは異なり、顕著な乳汁分泌活性を持っています。
下垂体前葉は、脂肪移動作用を持つ多くのペプチドおよびタンパク質物質を合成します。特に、ACTH、STH、TSHなどの下垂体刺激ホルモンは脂肪親和作用を有します。近年、特にβ-およびγ-脂肪親和性ホルモン(LPG)が注目されています。β-LPGの生物学的特性は最も詳細に研究されており、脂肪親和作用に加えて、メラノサイト刺激作用、副腎皮質刺激ホルモン刺激作用、低カルシウム血症作用、そしてインスリン様作用も示しています。
現在、豚と牛の脂肪親和性ホルモンであるヒツジLPG(90アミノ酸残基)の一次構造が解読されています。このホルモンには種特異性がありますが、β-LPGの中心領域の構造は種間で共通しており、それがホルモンの生物学的特性を決定づけています。この領域の断片の一つが、α-MSH、β-MSH、ACTH、β-LPGの構造に見られます。これらのホルモンは進化の過程で同一の前駆体から生じたと考えられています。γ-LPGはβ-LPGよりも脂肪親和性が弱いです。
メラノサイト刺激ホルモン
下垂体中葉で合成されるこのホルモンは、その生物学的機能において、皮膚色素メラニンの生合成を刺激し、両生類の皮膚における色素細胞メラノサイトの大きさと数の増加を促進します。MSHのこれらの特性は、ホルモンの生物学的試験に利用されています。このホルモンには、α-MSHとβ-MSHの2種類があります。α-MSHには種特異性が存在せず、すべての哺乳類で同じ化学構造を持つことが示されています。その分子は、13個のアミノ酸残基からなるペプチド鎖です。一方、β-MSHには種特異性があり、動物ごとに構造が異なります。ほとんどの哺乳類において、β-MSH分子は18個のアミノ酸残基で構成されていますが、ヒトにおいてのみ、アミノ酸末端から4個のアミノ酸残基が延長されています。α-MSHには副腎皮質刺激作用があり、動物およびヒトの行動への影響が実証されていることに留意する必要があります。
オキシトシンとバソプレシン
視床下部で合成されるバソプレシンとオキシトシンは、下垂体後葉に蓄積します。バソプレシンは視索上核のニューロンに、オキシトシンは室傍核に蓄積されます。その後、これらは下垂体へと移行します。ここで注目すべきは、バソプレシンホルモンの前駆体がまず視床下部で合成されるということです。同時に、ニューロフィシン1型と2型というタンパク質がそこで産生されます。前者はオキシトシンと結合し、後者はバソプレシンと結合します。これらの複合体は、神経分泌顆粒の形で細胞質内を軸索に沿って移動し、下垂体後葉に到達します。そこで神経線維は血管壁で終結し、顆粒の内容物が血液中に放出されます。バソプレシンとオキシトシンは、アミノ酸配列が完全に確立された最初の下垂体ホルモンです。化学構造的には、ジスルフィド結合を1つ持つノナペプチドです。
検討されているホルモンは、様々な生物学的作用を有します。膜を介した水と塩分の輸送を促進し、昇圧作用を有し、出産時の子宮平滑筋の収縮を促進し、乳腺からの分泌を増加させます。バソプレシンはオキシトシンよりも高い抗利尿作用を有し、オキシトシンは子宮と乳腺への作用がより強いことに留意する必要があります。バソプレシン分泌の主な調節因子は水分摂取です。尿細管において、バソプレシンは細胞膜上の受容体に結合し、その後、受容体内のアデニル酸シクラーゼという酵素を活性化します。ホルモンと受容体の結合と生物学的作用は、分子の様々な部位によって制御されています。
視床下部を介して神経系全体と繋がる下垂体は、内分泌系を機能的に統合し、体内環境の恒常性(ホメオスタシス)の維持に関与しています。内分泌系においては、下垂体前葉と「標的」腺(甲状腺、副腎皮質、性腺)間のフィードバック原理に基づいて恒常性調節が行われます。「標的」腺で産生されるホルモンが過剰になると抑制され、不足すると対応する刺激ホルモンの分泌と放出が促進されます。このフィードバック系には視床下部が含まれており、視床下部には「標的」腺のホルモンに感受性のある受容体領域が存在します。視床下部受容体は、血中を循環するホルモンに特異的に結合し、ホルモン濃度に応じて反応を変化させることで、その作用を対応する視床下部中枢に伝達します。視床下部中枢は下垂体の働きを調整し、視床下部から下垂体前葉刺激ホルモンを放出します。したがって、視床下部は神経内分泌脳とみなされるべきです。