
エシェリキオース(エシェリキア属、大腸菌)
最後に見直したもの: 06.07.2025

すべてのiLiveコンテンツは、可能な限り事実上の正確さを保証するために医学的にレビューまたは事実確認されています。
厳格な調達ガイドラインがあり、評判の良いメディアサイト、学術研究機関、そして可能であれば医学的に査読された研究のみにリンクしています。 かっこ内の数字([1]、[2]など)は、これらの研究へのクリック可能なリンクです。
当社のコンテンツのいずれかが不正確、期限切れ、またはその他の疑問があると思われる場合は、それを選択してCtrl + Enterキーを押してください。
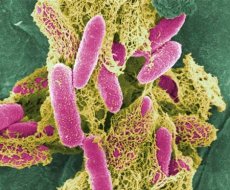
エシェリヒア属の代表的な細菌である大腸菌(E. coli)は、1885年にT.エシェリヒによって初めて発見され、この属は彼の名にちなんで命名されました。この属の主な特徴は、周毛性(運動性がない)、乳糖を発酵させて酸とガスを生成する(乳糖陰性)、クエン酸を含む飢餓培地では生育しない、フォーゲス・プロスカウアー反応は陰性、MR試験は陽性、フェニルアラニンデアミナーゼを持たず、KCNを含む培地では生育しない、DNA中のG + C含有量は50~51モル%です。
大腸菌属には少なくとも7種が含まれますが、医学上特に重要なのは大腸菌であり、特にヒト疾患を引き起こす変異株が重要です。大腸菌は、腸管外疾患を引き起こすものと急性腸疾患(AID)を引き起こすものの2つの主要なグループに分けられます。腸管外疾患の代表例は、以下の3つの病理学的グループに分類されます。
- 髄膜炎(MENEC - 髄膜炎大腸菌)
- 敗血症(SEPEC - 敗血症大腸菌)および
- 尿路病原性(UPEC - 尿路病原性大腸菌)。
急性呼吸器感染症を引き起こす大腸菌の変異株は、当初、腸管毒素原性大腸菌(ETEC)、腸管侵襲性大腸菌(EIEC)、腸管病原性大腸菌(EPEC)、腸管出血性大腸菌(EHEC)の4つのカテゴリーに分類されていました。その後、腸管凝集性大腸菌(EAEC)とびまん性凝集性大腸菌(DAEC)の2つのカテゴリーが新たに特定されました。
さらに、大腸菌は、水、特に飲料水や食品の糞便汚染の程度を示す指標として国際基準で使用されています。
大腸菌の標準株(E. coli K-12)は、細菌の遺伝学を研究するために世界中の多くの国の研究室で広く使用されています。
 [
[ 形態学
大腸菌は通性嫌気性菌で、従来の栄養培地でよく生育します。寒天培地上のコロニーは丸く、凸状で、半透明です。培養液上では、拡散した濁りを呈します。生育の最適温度は37℃で、10~45℃の範囲で生育します。最適pHは7.2~7.5です。乳糖を分解する大腸菌のコロニーは、すべての鑑別診断培地上で指示薬の色(エンド培地では金属光沢のある濃い赤色)を呈します。
生化学的性質
多くの場合、大腸菌は、グルコース、ラクトース、マンニトール、アラビノース、ガラクトース、そして時にはスクロースやその他の炭水化物を発酵させて酸とガスを生成する能力があります。インドールを生成しますが、通常はH2Sを生成しません。硝酸塩を亜硝酸塩に還元し、ゼラチンを液化せず、クエン酸を含む飢餓培地では増殖せず、MR反応で陽性反応を示し、Voges-Proskauer反応で陰性反応を示します。これらの特徴から、多くの疾患(赤痢、腸チフス、サルモネラ症など)の病原体と容易に区別できます。しかしながら、病原性大腸菌は、培養特性や生化学的特性において非病原性大腸菌と差がないことが非常に多くあります。
大腸菌の病原性因子
大腸菌がさまざまな病気を引き起こす能力は、以下の病原性因子の存在によるものです。
接着因子と定着因子。これらは組織細胞への接着と定着に必要です。定着因子には3つの変異体が発見されています。a) CFA/I-CFA/VI(定着因子) - 線毛構造を有します。b) EAF(腸管病原性大腸菌接着因子) - インチミン - 外膜タンパク質で、eaeA遺伝子によってコードされています。E. coli 4およびEHECに存在し、細菌がHep-2細胞に接着する能力によって検出されます。c) Henle-407接着因子 - 線毛構造で、細菌がHenle-407細胞に接着する能力によって検出されます。これらはすべてプラスミド遺伝子によってコードされています。これらに加えて、細菌性リポ多糖類を含む他の定着因子も報告されています。
侵入因子。例えば、EIECやEHECは、これらの因子の助けを借りて腸管上皮細胞に侵入し、そこで増殖して細胞を破壊します。侵入因子の役割は、外膜タンパク質によって担われます。
外毒素。病原性大腸菌には、膜を損傷する外毒素(溶血素)、タンパク質合成を阻害する外毒素(志賀毒素)、二次メッセンジャー(伝達物質)を活性化する外毒素(CNF、ST、CT、CLTD、EAST など)が含まれていることが分かっています。
溶血素は、大腸菌を含む様々な病原体によって産生されます。溶血素は孔形成毒素であり、標的細胞膜に結合して孔を形成し、そこから小分子やイオンが出入りすることで細胞死と赤血球溶解を引き起こします。
志賀毒素(STX)は、赤痢菌(Shigella dysenteriae)で初めて発見され、その後、EHEC(腸管出血性大腸菌)で類似の毒素(志賀様毒素)が発見されました。この毒素(N-グリコシダーゼ)は、28S rRNAと相互作用することでタンパク質合成を阻害し、細胞死(細胞毒素)を引き起こします。志賀様毒素には、STX-1とSTX-2の2種類があります。STX-1は抗原特性において志賀毒素とほぼ同一ですが、STX-2は抗原特性において志賀毒素と異なり、STX-1とは異なり、抗血清によって中和されません。大腸菌におけるSTX-1およびSTX-2細胞毒素の合成は、中等度の変換プロファージ9331(STX-1)および933W(STX-2)の遺伝子によって制御されています。
- 毒素 L (熱不安定毒素) は ADP-リボシルトランスフェラーゼであり、G タンパク質に結合して下痢を引き起こします。
- ST 毒素 (耐熱性毒素) はグアニル酸シクラーゼ受容体と相互作用し、その活性を刺激して下痢を引き起こします。
- CNF(細胞傷害性壊死因子)は、いわゆるRhoGタンパク質に損傷を与える脱アミダーゼタンパク質です。この毒素は、尿路感染症を引き起こすUPECに含まれています。
- CLTD毒素は細胞致死性崩壊毒素である。その作用機序は十分に解明されていない。
- EAST 毒素は、腸管凝集性大腸菌 (EAEC) の熱安定性毒素であり、おそらく熱安定性毒素 (ST) に類似しています。
エンドトキシンはリポ多糖類です。細菌の抗原特異性(糖の側鎖の繰り返しによって決定)とコロニーの形状(側鎖の喪失によりSコロニーがRコロニーに変化する)を決定します。
このように、大腸菌の病原性因子は、宿主細胞の染色体遺伝子だけでなく、プラスミドや溶原性転換ファージによって導入された遺伝子によっても制御されています。これらすべては、プラスミドや溶原性転換ファージが大腸菌間で拡散することで、病原性変異体が出現する可能性があることを示唆しています。以下は、急性呼吸器感染症を引き起こす大腸菌の4つのカテゴリーについて簡単に説明します。最近特定されたDAECおよびEAECのカテゴリーに関する情報は、入手可能な情報源には見つかりませんでした。
ETECには17の血清群が含まれます。CFA型の繊毛構造の接着因子および定着因子とエンテロトキシン(LTまたはST、あるいはその両方)は、同じプラスミドによってコードされています。これらは絨毛に損傷を与えることなく定着します。エンテロトキシンは水塩代謝の障害を引き起こします。感染部位は小腸です。感染量は10~10個です。この疾患はコレラ様下痢として進行します。流行は水系で、食品媒介はまれです。1~3歳の幼児と成人が罹患します。
EIECには9つの血清群が含まれ、病原性は腸粘膜上皮細胞に浸透して増殖し、細胞を破壊する能力に関連しています。これらの特性は、染色体遺伝子に加えて、プラスミド遺伝子(140 MD)によってコード化されています。プラスミドは、侵入を決定する外膜タンパク質の合成をコード化します。プラスミド自体とそれがコード化するタンパク質はどちらも赤痢病原体のものと関連しており、これがEIECと赤痢菌の類似性を説明しています。感染量は10s細胞です。プロセスの局在は、下部回腸と大腸です。病気は赤痢として進行します。最初は水様性下痢、次に大腸炎症候群です。1.5〜2歳の子供、青年、成人が病気になります。発生の種類 - 食品、水。
疫学
大腸菌は、あらゆる哺乳類、鳥類、爬虫類、魚類の腸管に生息する正常な微生物叢の代表です。したがって、どの大腸菌の変異体がなぜ大腸菌症を引き起こすのかという疑問を解明するためには、抗原構造を研究し、病原性血清変異体を同定するために必要な血清学的分類を開発し、それらがどのような病原性因子を有するか、すなわちなぜ様々な形態の大腸菌症を引き起こすことができるのかを解明する必要がありました。
大腸菌には、O抗原変異体(O1~O171)が171種、H抗原変異体(H1~H57)、および表面(莢膜)K抗原変異体が90種存在します。しかし、実際には、O抗原群は164種、H抗原血清型は55種存在します。これは、以前のO:H血清群の一部が大腸菌種から除外された一方で、O抗原とH抗原の序数は変更されていないためです。下痢原性大腸菌の抗原特性には、O抗原番号とH抗原番号が含まれます。例えば、O抗原は特定の血清群に属していることを示し、H抗原はその血清型変異体です。さらに、O抗原およびH抗原のより詳細な研究により、いわゆるO因子抗原およびH抗原、すなわちH2a、H2b、H2c、あるいはO20、O20a、O20abなどの抗原サブバリアントが明らかになりました。下痢原性大腸菌のリストには、合計43のO血清群と57のOH血清群が含まれています。このリストは常に新しい血清群で補完されています。
症状
このグループには、9つのクラス1血清群と4つのクラス2血清群が含まれます。クラス1血清群には、EAF型の接着および定着因子の合成を制御するプラスミド(60MDa)があります。これは外膜に局在するタンパク質によって表され、細菌がHEp-2細胞に付着する能力によって検出されます。タンパク質の分子量は94kDaです。クラス2血清群にはこのプラスミドがありません。その病原性は他のいくつかの要因によるものです。両方のクラスの一部の株は、STXを合成できることがわかっています。4は腸管上皮細胞の細胞膜に定着し、上皮表面に損傷を与え、びらんと中程度の炎症を形成します。感染量は105〜10 12個の細胞です。このプロセスは小腸に局在します。この病気は水様性下痢と重度の脱水症状が特徴です。主に生後1年目の子供が病気になります。感染経路は家庭内での接触感染ですが、食品媒介感染は稀です。
血清群 EIEC および 4 は院内感染の発生の最も一般的な原因です。
EHECは細胞毒素STX-1およびSTX-2を産生します。ヒトにおいて出血性大腸炎を引き起こし、溶血性尿毒症や血栓性血小板減少性紫斑病などの重篤な合併症を引き起こします。これらの毒素は小血管の内皮細胞を破壊します。血栓の形成とフィブリン沈着は、血流障害、出血、虚血、細胞壁の壊死につながります。尿毒症性溶血症候群は致命的となる可能性があります。EHECには多くの血清型(-150)がありますが、疫学的に主要な役割を果たしているのは、STXを産生する大腸菌O157-H7とその鞭毛変異株であるO157:NMです。これらの菌株は、細胞毒素の1つだけを分泌することも、両方を同時に分泌することもできます。大腸菌O157:H7を含む腸管出血性大腸菌(EHEC)血清型の自然宿主は、牛と羊であると考えられています。最も一般的な感染経路は食品(肉、特にひき肉、牛乳)です。大腸菌O157:H7は、有害な因子に対して非常に強い耐性を持っています。これが、様々な食品中での生存と増殖に寄与しています。感染は家庭用品との接触によって起こる可能性があります。発症は急性で、腸の痙攣が起こり、その後下痢が起こります。最初は水様性で、その後血便が続きます。子供も大人も発症します。感染者は感染性があります。
臨床診断
これは、病原体の純培養の分離と同定、およびPCRを用いた毒素検査に基づいています。大腸菌症の原因物質は、多価OK血清のセットと、特定の抗原に対する抗体のみを含む吸着血清のセットを用いて同定されます。角結膜テストはEIECの同定に使用できます。EIECの一部は不動性で、ラクトースとサリシンを発酵しません。大腸菌O157:H7の同定は、ソルビトールを発酵できないことで容易になります(ラクトースの代わりにソルビトールを含むエンド培地を使用します)。しかし、OKZ病原体(すべてのカテゴリ)の同定と区別には、PCRテストシステムを使用するのが最適です。必要に応じて、分離された病原体の抗生物質に対する感受性を決定します。