
エシェリキア属(エシェリヒア属、大腸菌)
最後に見直したもの: 23.04.2024

すべてのiLiveコンテンツは、可能な限り事実上の正確さを保証するために医学的にレビューまたは事実確認されています。
厳格な調達ガイドラインがあり、評判の良いメディアサイト、学術研究機関、そして可能であれば医学的に査読された研究のみにリンクしています。 かっこ内の数字([1]、[2]など)は、これらの研究へのクリック可能なリンクです。
当社のコンテンツのいずれかが不正確、期限切れ、またはその他の疑問があると思われる場合は、それを選択してCtrl + Enterキーを押してください。
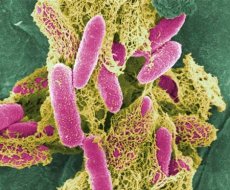
エシェリヒア属の主な代表 - 大腸菌 - 最初の1885年に発見された、T.大腸菌、その名誉に細菌のこの属は、その名前を得ました。この種の主な機能:周毛(または固定)発酵ラクトースが増殖しないクエン酸飢餓培地中の酸及びガス(又はlactosonegative)を形成する、フォゲス-Proskauer反応はfenilalanindezaminazyがない培地上で増殖有する、MR試験が陽性であると負でありますKCNでは、DNA中のG + C含量は50〜51モル%である。
エシェリヒア属には少なくとも7種が含まれる。医学において特に重要なのは、大腸菌(E.coli)の形態、特にヒト疾患を引き起こすその変異体の形態である。彼らは2つの主要なグループに分かれています:腸内の病気(OKZ)の腸内病気と病原体を引き起こします。最初の代表者は3つの病理グループに分けられます:
- 髄膜炎(MENEC - 髄膜炎;大腸菌);
- 敗血症(SEPEC-敗血症E.coli)および
- 泌尿器病原性(UPEC - 泌尿器病原性大腸菌)。
次に、OKZを引き起こす大腸菌の変異体は、最初に以下の4つのカテゴリーに分類された:腸溶性原性大腸菌(ETEC); 腸内侵襲性大腸菌(EIEC); 腸内病原性大腸菌(EPEC)および腸管出血性大腸菌(EHEC)。続いて、腸内凝集性大腸菌(EAEC)および拡散性凝集性大腸菌(DAEC)の2つのさらなるカテゴリーが同定された。
さらに、大腸菌は、水、特に飲料水および食品の糞便汚染の程度の指標として、国際基準で使用されている。
大腸菌の標準株(大腸菌K-12)は、細菌の遺伝学を研究するために世界の多くの国の研究所で広く使用されている。
 [
[形態学
大腸菌 - 通性嫌気性、通常の栄養培地でよく生育します - 寒天のコロニーは丸い凸状、半透明です。濁った濁りの形のブロス上での生育。37°Cの増殖に最適な温度は10〜45°Cの範囲で増加し、最適pHは7.2〜7.5である。コロニーE.coliの全ての示差的診断環境について。ラクトースを分解する。インジケータの色で塗装されています(金属光沢のある遠赤外線の環境で)。
生化学的性質
大腸菌は、大抵の場合、グルコース、ラクトース、マンニトール、アラビノース、ガラクトース、時にはスクロースおよび他のいくつかの炭水化物を生成するために、以下の炭水化物を発酵させることができる。インドールを形成する。原則として、H 2 Sを形成しない。硝酸塩を硝酸塩に戻し、ゼラチンを希釈せず、クエン酸塩を含む飢餓培地で増殖せず、MRと陽性反応を生じ、Foges-Proskauerと陰性反応を示す。これらの兆候によって、それは多くの病気(赤痢、腸チフス、サルモネラなど)の病原体と容易に区別することができます。しかし、病原性大腸菌は、培養および生化学的特性の両方に関して、非病原性のものとしばしば異なるものではない。
エシェリヒア・コリの病原性因子
種々の疾患を引き起こす大腸菌の能力は、病原性の以下の因子の存在に起因する:
接着とコロニー形成の要因。それらは、組織細胞への付着およびそれらのコロニー形成に必要である。コロニー形成因子の3つの変種が見いだされた:a)CFA / I-CFA / VI(英国植民地化因子) - それらは線維構造を有する。b)EAF(英語の腸内病原性大腸菌接着因子) - インチミンは外膜タンパク質であり、eAeA遺伝子によってコードされる。Hep-2の細胞に付着する細菌の能力によって明らかにされた4およびEHECに見いだされる; c)付着Henle-407 - 線毛構造は、細菌がHenle-407細胞に付着する能力によって検出される。それらはすべてプラスミド遺伝子によってコードされる。これらに加えて、コロニー形成の他の因子が記載されており、その役割において、細菌性リポ多糖類もまた作用し得る。
侵略の要因。彼らの助けを借りて、例えば、EIECとEHECは、腸の上皮細胞に浸透し、増殖し、その破壊を引き起こす。浸潤因子の役割は外膜タンパク質によって行われる。
外毒素。病原性大腸菌で外毒素者らは、第二メッセンジャー活性化タンパク質合成(志賀毒素)を阻害損傷膜(溶血素)、(ENGLメッセンジャー - コヒーレント) - CNF毒素、ST、CT、CLTD、東。
ヘモリシンは、大腸菌を含む異なる病原体を産生する。ヘモリシンは細孔形成毒素である。最初に標的細胞の膜と接触し、次いで小さな分子およびイオンが出入りする孔を形成し、細胞死および赤血球溶解を導く。
志賀毒素(STX)は志賀赤痢菌で最初に発見した後、同様の毒素(shigapodobny毒素)はEHECに見出されました。毒素(N-グリコシダーゼ)は、28S rRNAと相互作用することによってタンパク質合成をブロックし、その結果、細胞が死滅する(細胞毒素)。滋養物様の毒素には、STX-1とSTX-2の2種類があります。STX-1がそれに対する抗血清によって中和されていないとは異なり、志賀毒素とほぼ同じSTX-1抗原特性は、STX-2は、志賀毒素抗原特性とは異なり、。STX-STX-1及び2は、大腸菌遺伝子に制御される合成の細胞毒素は、変換プロファージ9331(STX-1)および933W(STX-2)中程度。
- 毒素L(熱不安定性毒素)-ADP-リボシルトランスフェラーゼ; Gタンパク質への結合は、下痢を引き起こす。
- グアニル酸シクラーゼの受容体と相互作用する毒素ST(熱安定性毒素)は、その活性を刺激し、下痢を引き起こす。
- CNF(細胞傷害性壊死因子) - タンパク質デアミダーゼは、いわゆるRhoGタンパク質に損傷を与えます。この毒素はUPECに認められ、尿路感染を引き起こします。
- CLTD-毒素は、細胞傷害性緩和毒素である。行動の仕組みはあまり理解されていない。
- 毒素EASTは、おそらく熱安定性毒素(ST)に類似した腸内凝集性大腸菌(EAEC)の熱安定性毒素である。
エンドトキシン - リポ多糖類。それらは細菌の抗原特異性(糖の反復側鎖によって決定される)およびコロニーの形状(側鎖の喪失はRコロニーにおけるSコロニーの形質転換を導く)を決定する。
したがって、大腸菌の病原性因子は、宿主細胞の染色体遺伝子だけでなく、プラスミドによって、または中程度に変換するファージによって導入された遺伝子によっても制御される。この全ては、プラスミドおよびその中の中等度のファージの広がりの結果としてのE.coliの病原性変異体の可能性を示している。以下は、OKZを引き起こす大腸菌の4つのカテゴリーの簡単な説明である。私たちが入手可能な情報源のDAECとEAECの最近特定されたカテゴリーに関する情報は見つかりませんでした。
ETECは17の血清群を含む。CFA型およびエンテロトキシン(LTまたはST、またはその両方)の線状構造の接着およびコロニー形成因子は、同じプラスミド(プラスミド)によってコードされる。それらを損なうことなく絨毛を植え付ける。腸毒素は水 - 塩代謝の違反を引き起こす。プロセスの局在化は、小腸の領域である。108-1010細胞の感染用量。この疾患は、コレラ様の下痢のタイプに応じて進行する。流行のタイプ - 水、食べ物の頻度は低いです。1歳から3歳までの子供と成人が病気です。
EIECは9つの血清群を含み、病原性は腸粘膜の上皮細胞に浸透しそれらの中で増殖してその破壊を引き起こす能力に関連する。これらの特性は、染色体遺伝子に加えて、プラスミド遺伝子(140MD)によってコードされる。プラスミドは、浸潤を決定する外膜タンパク質の合成をコードする。プラスミド自体およびそれがコードするタンパク質は、赤痢の原因物質のものと関連しており、EIECと赤痢菌との間の類似性を説明している。10sの細胞を感染させる用量。プロセスの局在 - 下部回腸および大腸。この疾患は赤痢のタイプ、すなわち最初の水様の下痢、次いで大腸炎症候群に応じて進行する。子供は、2〜5年間、青年と成人に病気です。アウトブレイクの種類 - 食糧、水。
疫学
大腸菌は、哺乳類、鳥類、爬虫類および魚類の腸管の正常な微生物叢のメンバーです。したがって、質問を明確にするために、大腸菌や原因のオプションはehsherihiozom、抗原構造を研究するために必要な理由は、病原性serovariantamiの識別のために必要な血清学的分類を開発し、彼らはすなわち、病原性因子を持っているかを調べるものです。E.は、何らかの理由で、彼らはehsherihiozomの様々な形態を引き起こす可能性があります。
大腸菌では、見つかった171の変異体O-抗原(01から0171まで)、57-Hは、抗原(H1-H57)および90個の変異体表面(カプセル)K-抗原を実施例。Hの血清群種大腸菌から除外するが、O及びH抗原のシリアル番号は変化しないままであった:前の0の一部は、しかし、実際には164基は、O抗原及びH抗原の55 serovariantamiであります 下痢原性大腸菌の抗原特性には、O抗原およびH抗原の数、例えば055:116; 0157:H7; O-抗原は特定の血清群に属することを意味し、H-抗原はその血清型変異体である。さらに、識別OおよびH抗原の深さの調査でとても例えばすなわち、抗原性subvariants、因子OとH抗原と呼ばれる.. H2A、N2、またはH2C 020 O20a、O20ab等...合計で、43のO-血清型および57のOH-血清型が、下痢性大腸菌のリストに含まれる。このリストには、すべての新しい血清型が補充されています。
症状
群は、クラス1の9つの血清群およびクラス2の4つの血清群を含む。クラス1の血清群は、接着因子の合成およびEAF型のコロニー形成を制御するプラスミド(60MD)を有する。それは、外膜に局在するタンパク質によって表され、HEp-2の細胞に付着する細菌の能力によって同定される。このタンパク質の質量は94kDである。クラス2の血清群では、このプラスミドは存在せず、その病原性は他のいくつかの要因に起因する。両方のクラスのいくつかの系統4は、STXを合成する能力を有する。4は腸細胞の形質膜に定着し、浸食の形成および中程度の炎症を伴う上皮の表面の損傷を引き起こす。105-10 12細胞の感染用量。この過程は小腸に局在する。この疾患は、水様の下痢および顕著な脱水によって特徴付けられる。人生の最初の年のほとんどの子供たちは病気です。感染の方法は接触家庭で、食べる頻度は少なくなります。
血清群EIECおよび4は、院内感染の最も頻繁な原因である。
細胞毒STX-1およびSTX-2を産生する。人々は、溶血性尿毒症および血栓性血小板減少性紫斑病の形で重篤な合併症を伴う出血性大腸炎を引き起こす。毒素は小血管の内皮細胞を破壊する。血餅の形成およびフィブリンの喪失は、細胞壁の血流、出血、虚血および壊死の侵害につながる。不穏な溶血性症候群は死に至ることがある。EHECは多くの血清型(-150)に代表されるが、主要な疫学的役割は大腸菌O157-H7およびそのEAフリー0157:NM非突然変異体突然変異体である。これらの菌株は、細胞毒素のうちの1つ、またはその両方を放出することができる。大腸菌O157:H7を含むENOV血清型の天然のリザーバは、ウシおよびヒツジであると考えられている。最も頻繁な感染経路は食物(肉、特に細かい肉、牛乳)です。大腸菌O157:H7は、不利な要因に対して異常に耐性がある。これは、異なる製品における生存と生殖に寄与する。接触や家庭による汚染の可能性があります。病気の発症は急性です:腸のけいれん、その後下痢、最初は水分、次に血液。子供と大人は病気です。病気の人は伝染性があります。
検査室の診断
それは、病原体の純粋な培養物の単離およびその同定ならびにPCRによる毒素の試験に基づいている。エスケイキシスの原因物質は、多価OK血清のセットおよび特定の抗原のみに対する抗体を含む吸着血清のセットで同定される。EIECを識別するために、角膜活性試料を使用することができる。一部のEIECの代表者は不動であり、乳糖とサリシンを発酵させない。大腸菌O157:H7の同定は、ソルビトールを発酵できないことによって助長される(乳糖の代わりにソルビトールを用いてEndo培地を用いる)。しかし、OKZ(すべてのカテゴリー)の病原体を同定および分化させるために、PCR試験システムを使用することが最善です。必要であれば、選択された病原体が抗生物質に対する感受性を決定する。