
卵巣
最後に見直したもの: 23.04.2024

すべてのiLiveコンテンツは、可能な限り事実上の正確さを保証するために医学的にレビューまたは事実確認されています。
厳格な調達ガイドラインがあり、評判の良いメディアサイト、学術研究機関、そして可能であれば医学的に査読された研究のみにリンクしています。 かっこ内の数字([1]、[2]など)は、これらの研究へのクリック可能なリンクです。
当社のコンテンツのいずれかが不正確、期限切れ、またはその他の疑問があると思われる場合は、それを選択してCtrl + Enterキーを押してください。
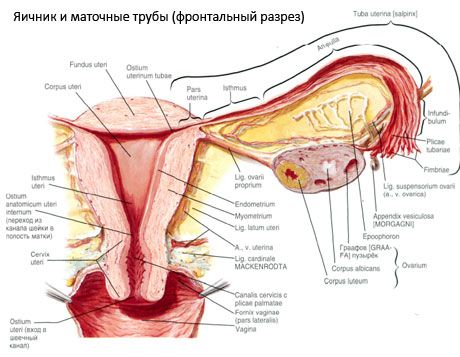
卵巣(卵管、ギリシャの卵胞子) - 子宮の広い靭帯の後ろにある小さな骨盤の腔に位置する臓器、女性の生殖腺。卵巣は、女性の性細胞(胚珠)、ならびに血液およびリンパに入る女性の性ホルモンを発生させ、成熟させる。卵巣は卵形をしており、前後方向にやや平らになっています。卵巣の色はピンク色です。出産した女性の卵巣の表面には、黄色い体の排卵や変形の跡が見られます。内側表面(顔貌のmedialisさ:2 cmまでの卵巣は、2つの自由表面を区別している - 8.5グラムの卵巣寸法は:. 2.5〜5.5センチメートルの長さ、1.5〜3.0センチメートル幅、厚さ、卵巣重量であるました卵巣窩 - )骨盤腔、部分的に覆われた卵管、及び側面(顔貌がlateralis)、隣接する骨盤側壁、軽度の深化の方を向いています。この中心窩は、上部および子宮の腹膜外腸骨血管と下部の咬合動脈との間のコーナーに位置する。卵巣の背後には、対応する辺の尿管が上から下に通過する。
腸間膜領域(マーゴのmesovaricus)で、腹膜の短いクリップ倍(腸間膜卵巣)によって子宮広間膜の後ろの葉に-卵巣表面が凸面フリー(後)端(マーゴの公文書)で、前方に移動します。卵巣は、動脈および神経位置静脈およびリンパ管を含み、それを通して卵巣ゲート(へそのovarii)、 -体の最前線に溝付きの凹部です。我々また、単離された卵巣両端:丸みを帯びた上部管状端(extremitasのtubaria)、卵管に面し、そして母(extremitasのutenna)の下端、子宮に結合された自身の束卵巣(LIG ovarii proprium)。母親から6ミリメートルの周りのラウンドコードでこのバンドルには、広範な靱帯の2枚の間に位置し、横方向の角卵巣、子宮、の終わりです。靱帯装置卵巣によっても関する靭帯podveshivayaschaya卵巣卵巣および卵巣を含む容器と繊維の繊維束で骨盤の上部壁から延びる腹膜のひだである(lig.suspensorium ovarii)を、。卵巣固定短い腸間膜卵巣腸間膜縁に子宮の尖広い靱帯から延びる、duplikatury腹膜を表す(mesovarium)。卵巣自体は腹膜に覆われていません。子宮管の最大の卵巣線毛は、卵巣の管末端に付着する。卵巣のトポグラフィは、子宮の位置、その大きさ(妊娠中)に依存する。卵巣は、骨盤腔の非常に可動性の器官を指す。
 [
[卵巣血管および神経
卵巣への血液供給は、aaによるものです。et vv。卵巣子宮および子宮頸。腎動脈 - 卵巣動脈(Aaとovaricae dextraらシニストラ)の両方が右が最も頻繁に大動脈および左由来する、ちょうど腎動脈の下の大動脈の正面表面から延びています。前面の腰の筋肉の上にダウンして横方向に行く、卵巣動脈は(彼に小枝を与える)各フロント尿管、外腸骨血管、および卵巣の提靭帯にここに配置されている骨盤腔に入る境界線と交差します。内側以下、卵巣動脈は、それに分岐を与えて、卵管の下子宮広間膜のシートの間に通過し、 - 卵巣の腸間膜に、卵巣の門に入る。
卵巣動脈の枝は、子宮動脈の卵巣枝で広く吻合されている。卵巣からの静脈流出は、卵巣門の領域に位置する卵巣静脈叢で主に行われる。したがって、血液の流出は、子宮と卵巣の静脈を通る2つの方向に進む。右卵巣静脈には弁があり、下部大静脈に通じている。左卵巣静脈は左腎静脈に流入し、そこには弁がない。
卵巣からのリンパ流出は、特に臓器ゲートの領域に豊富なリンパ管を通して起こり、舌下リンパ叢は隔離される。次いで、リンパは、卵巣リンパ管に沿って大動脈リンパ節に回される。
卵巣の収縮
交感神経 - 腹腔(太陽)、上編組および下胃の叢からの節後繊維によって提供される。副交感神経 - 内部の仙骨神経による。
卵巣の構造
表面は、卵巣胚上皮の単層で覆われています。その下には緻密な結合組織エンベロープ(tunica albuginea)があります。弾性繊維に富む卵巣形態支質(ストロマovarii)の結合組織。それは外側及び内側の層によって分割される物質卵巣実質。髄質(髄質ovarii)と呼ばれ、ゲートに近い卵巣の中心にある内層、、、。ゆるい結合組織のこの層には、多数の血液およびリンパ管および神経が存在する。卵巣の外層-皮質(皮質ovarii)密。これは熟成配置されている結合組織のたくさん持っている一次卵胞(小節ovarici primarii)、二次(バブル)卵胞、(小節ovarici secundarii、s.vesiculosi)をウェル気泡graafovy成熟卵胞(小節ovarici maturis)、およびイエローおよびatretic体。
各卵胞には、女性の生殖卵子または卵母細胞(卵母細胞)がある。丸み150ミクロンまで、卵母細胞の直径は、核、細胞小器官に加えて、タンパク質 - 脂質封入体(卵黄)があり、細胞質の大量、卵の供給のために必要なグリコーゲンを含有します。その卵母細胞の供給は、通常、排卵後12〜24時間以内に消費される。受精が起こらなければ、卵は死ぬ。
ヒト卵巣は2つのカバー膜を有する。内部には、卵母細胞の細胞質膜である細胞壁がある。細胞壁の外側には、卵を保護し、ホルモン形成機能(エストロゲン放出)を行う、いわゆる濾胞細胞の層がある。
子宮、管および卵巣の生理学的位置は、腹膜、靭帯および骨盤のセルロースを組み合わせた、懸濁、固定および支持装置によって提供される。サスペンション装置は、対形成された形態で表され、子宮の円形および幅広の靭帯、靭帯および垂下靱帯卵巣を含む。子宮の広い靱帯、卵巣の自身のおよび吊り下げ靭帯は、子宮を中間位置に保つ。丸い靭帯は、子宮の底を前方に引き付け、その生理学的な傾斜を提供する。
固定装置は、小さな骨盤の中心に震えの位置を確保し、それを前後にシフトさせることを事実上不可能にする。しかし、靱帯装置はその下部の子宮から離れるので、様々な方向に子宮を傾けることが可能である。仙腸magochnyeのカーディナル、子宮および嚢胞-vesico-恥骨靱帯:定着ユニットによってルーズ骨盤組織に配置され、側面、前面及び後壁骨盤の下に子宮カードから延びるコードを含みます。
中胚葉に加えて、以下の卵巣靭帯が区別される:
- 卵巣の提靭帯は、以前voronkotazovayaとして指定します。これは、(Ovarica。のEt v)をその中に血液を延びて腹膜のひだであり、そして上部(チューブ)(外側および内側に共通の腸骨動脈を分割に)横骨盤壁、腰筋膜との間に張設卵巣リンパ管および神経卵巣の終わり。
- 卵巣自身の靭帯は、幅の広い子宮靭帯のシートの間を後ろのシートの近くに通し、卵巣の下端を子宮の側縁に接続する。子宮には、卵巣自身の靭帯が、子宮管の始まりと円形の靭帯の間に、後者から前後に取り付けられている。靱帯の厚さはrrです。子宮動脈の終末枝である卵巣;
- 虫垂 - 卵巣靭帯クレードは、腹膜の襞の形で、卵巣または右卵巣または子宮の広い靭帯に虫垂の隆起部の周りに延びる。靭帯は不安定で、女性の1/2〜1/3で観察されます。
支持装置は、骨盤底の筋肉および筋膜によって表され、下部、中間および上部(内側)の層に分割される。
最も強力なのは、肛門を引き上げる対の筋肉によって表される上(内)の筋肉層です。これは3つの方向(恥骨尾骨-、尾椎-ILIOとischiococcygeal筋肉)における骨盤への尾骨からファンアウトの筋肉の束で構成されています。この筋層は、骨盤の横隔膜とも呼ばれます。
筋肉の中間層は、交感神経、恥骨および坐骨の骨の間に位置する。筋肉の中間層 - 泌尿生殖器の横隔膜 - は骨盤口の前半分を占め、尿道と膣を通過する。その葉の間の前部には、尿道の外側括約筋を形成する筋肉梁があり、後部には、横方向に伸びる筋束、会陰の横方向の筋肉がある。
以下の下側(外側)層骨盤底筋は、これらは、球海綿体、海綿含む図8のような形をしている場所の表面、から成る、坐骨海綿-、肛門の外部括約筋、浅会陰横筋。
卵巣の発生原性
成長および濾胞閉鎖のプロセスは、20週間の妊娠から始まり、少女の卵巣への送達の時間までに200万個の卵母細胞が残る。初夏の時までに、その数は30万に減少し、生殖器の全期間中に成熟し、500個以下の卵胞が排卵される。卵胞の初期成長はFSHの刺激に依存せず、限られており、閉鎖は迅速に起こる。ステロイドホルモンの代わりに、局所オートクリン/パラクリンペプチドが、原発性卵胞の成長および退縮の主な調節因子であると考えられている。毛包の成長および閉鎖のプロセスは、生理学的プロセスによって中断されないと考えられている。このプロセスは、子宮内の期間および更年期を含むすべての年齢で継続し、妊娠、排卵および無排卵によって中断される。特定のサイクルごとに卵胞の成長およびその数を引き起こすメカニズムはまだ明らかではない。
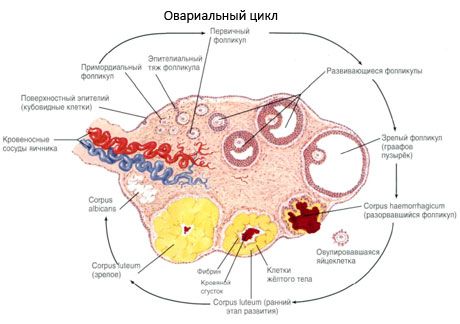
その発達において、小胞はいくつかの発達段階を経る。始原生殖細胞は、卵黄嚢の内胚葉由来であり、妊娠5〜6週目に胎児の生殖器領域に移動する。妊娠6〜8週から16〜20週まで続く迅速な有糸分裂の結果、肉芽腫細胞の薄い層に囲まれた胚の卵巣に最大6-7百万の卵母細胞が形成される。
前卵胞 - 卵母細胞は膜(Zona pellucida)に囲まれている。卵母細胞を取り囲む顆粒膜細胞は増殖し始め、それらの増殖はゴナドトロピンに依存し、エストロゲンのレベルと相関する。顆粒膜細胞はFSHの標的である。前胞の段階で、顆粒膜細胞は、アンドロゲンをエストラジオールに変換する主な酵素であるアロマターゼの活性を優先的に誘導する3つのクラスのステロイドを合成することができる。エストラジオールは、それ自体の受容体の数を増加させることができ、FSHとは無関係に顆粒膜細胞に直接分裂促進作用を与えると考えられている。これは芳香族化プロセスの活性化を含むFSHの効果を高めるパラクリン因子と考えられている。
FSH受容体は、卵胞の成長が始まるとすぐに顆粒膜細胞の膜上に現れる。FSHの減少または増加は、その受容体の数の変化につながる。FSHのこの作用は成長因子によって調節される。イオンチャネル、二次メッセンジャーの受容体チロシンキナーゼ、ホスホリパーゼシステム:FSHは、主規制FSHものの卵胞ステロイドでGタンパク質アデニル酸シクラーゼ系を介して作用する、プロセスは多くの要因が関与します。
卵胞の初期発生におけるアンドロゲンの役割は複雑である。顆粒膜細胞はアンドロゲン受容体を有する。彼らは、エストロゲンへのFSH誘発性の芳香族化のための基板のみではなく、低濃度で香料のプロセスを強化することができます。場合アンドロゲンのレベルは、エストロゲンおよびエストロゲンに変換することができない現像アンドロゲン、アンドロゲンでの5α-レダクターゼ、およびそれによって阻害アロマターゼ活性を介して変換する簡単な方法に全く芳香族化前胞状顆粒膜細胞が優先的に選択された経路を増加しません。このプロセスはまた、FSHおよび LH受容体の形成を阻害し、かくして卵胞の発達を停止させる。
高レベルのアンドロゲンを伴う卵胞の芳香化のプロセスは、退縮のプロセスを経る。卵胞の成長および発達は、アンドロゲンをエストロゲンに変換する能力に依存する。
FSHの存在下では、卵胞液の主要な物質はエストロゲンである。FSH - アンドロゲンがない場合。LHは、循環の途中まで卵胞液において正常である。できるだけ早くLHの顆粒膜細胞の有糸分裂活性の増加したレベルが減少する、及び変性変化は、血漿中より卵胞液中の卵胞ステロイドレベルの増加アンドロゲンレベルが表示され、機能的活性卵巣反映:顆粒と卵胞膜細胞。FSHの唯一の標的が顆粒膜細胞である場合、LHは多くの標的を有する - これらは細胞、間質および黄体細胞および顆粒膜細胞である。ステロイド生成に対する能力は、顆粒膜細胞およびテカ細胞の両方を有するが、アロマターゼ活性は顆粒膜細胞で優勢である。
LHに応答して、テカ細胞はアンドロゲンを産生し、次いで、FSH誘導性の芳香族化を介して、顆粒膜細胞によってエストロゲンに変換される。
卵胞の卵胞膜として細胞はLH受容体P450秒及び3ベータ - ヒドロキシステロイドデヒドロゲナーゼのための遺伝子を発現し始め、インスリン様成長因子(IGF-1)相乗LHで遺伝子の発現を増加させるが、ステロイドを刺激しません。
卵巣ステロイド生成は常にLH依存性である。卵胞が成長すると、現在の細胞は、コレステロールからアンドロゲンを形成するP450c17酵素を発現する。顆粒膜細胞はこの酵素を有さず、アンドロゲン由来のエストロゲンの産生において現在の細胞に依存している。ステロイド生成とは異なり、胞子形成はFSHに依存する。卵胞のようおよびエストロゲンのレベルを上げるアクション・フィードバック・メカニズムに入ってくる - FSHの産生を阻害、順番にアポトーシス(プログラム細胞死)を介してアロマターゼ活性包の減少につながると、最終的には卵胞閉鎖へ。
エストロゲンおよびFSHのフィードバック機構は、増殖を開始した卵胞の発達を阻害するが、支配的な卵胞を阻害しない。優性卵胞は、顆粒膜細胞の増殖およびエストロゲン中のアンドロゲンの芳香族化を支援するより多くのFSH受容体を含む。さらに、パラクリンおよびオートクリン経路は、卵胞の発生の重要な調整因子として働く。
オートクリン/パラクリン制御因子の不可欠な部分は、FSHの作用に応答して顆粒膜細胞によって合成され、卵胞液に入るペプチド(インヒビン、アクチビン、フォリスタチン)である。インヒビンはFSH分泌を減少させる。アクチビンは、下垂体からのFSHの放出を刺激し、卵巣におけるFSHの作用を増強する; フォリスタチンはおそらくはアクチビンの結合に起因するFSH活性を抑制する。排卵および黄色体の発生後、インヒビンはLHの制御下にある。
卵巣細胞の増殖および分化は、インスリン様増殖因子(IGE)の影響を受ける。IGF-1は顆粒膜細胞に作用し、環状アデノシン一リン酸(cAMP)、プロゲステロン、オキシトシン、プロテオグリカンおよびインヒビンの増加を引き起こす。
IGF-1は、テカ細胞に作用し、アンドロゲンの産生を増加させる。Teka細胞はまた、FSHによっても調節される腫瘍壊死因子(TNF)および上皮増殖因子(EGF)を産生する。
EGFは顆粒膜細胞の増殖を刺激する。IGF-2は卵胞液の増殖の主要因子であり、IGF-1、TNF-α、TNF-3およびEGFも検出した。
卵巣機能のパラクリンおよび/または自己分泌調節の違反は、排卵プロセスの障害および多嚢胞性卵巣の形成において役割を果たすようである。
卵胞が成長すると、卵胞液中のエストロゲンの含量が増加する。顆粒膜細胞の増加のピークにおいて、LHの受容体が現れ、顆粒膜細胞の黄体形成が起こり、プロゲステロン産生が増加する。従って、前排卵期の間に、エストロゲンの産生の増加はLH受容体の出現を引き起こし、LHは顆粒膜細胞の黄体形成およびプロゲステロン産生を引き起こす。プロゲステロンの増加はエストロゲンのレベルを低下させ、エストロゲンは明らかにサイクルの途中でFSHの第2のピークを引き起こす。
排卵は、LHのピークの10〜12時間後およびエストラジオールのピークの24〜36時間後に起こると考えられている。LHは、卵母細胞の減少、顆粒膜細胞の黄体形成、卵胞中のプロゲステロンおよびプロスタグランジンの合成を刺激すると考えられている。
プロゲステロンは、卵胞の壁を破裂させることに関与するプロスタグランジンと一緒にタンパク質分解酵素の活性を高める。プロゲステロン誘発FSHピークは、プラスミノーゲンをタンパク質分解酵素 - プラスミンに変換することによって卵母細胞を卵胞から排出させ、黄体期の正常な発達に十分な数のLH受容体を提供する。
ルテイン - 排卵、顆粒膜細胞が増加した後3日以内に、彼らは、顔料を充填特性液胞現れます。卵胞膜ルテイン細胞は卵胞膜および間質から分化および黄体の一部となります。非常に迅速に血管新生因子の影響下黄体に浸透毛細血管の発達であり、血管新生の改善にプロゲステロンとエストロゲンの生産を増加させました。活性ステロイドおよび黄体の寿命の持続時間は、LHのレベルによって決定されます。黄色の体は均一な細胞の実体ではありません。2種類の黄体細胞の添加、それは内皮細胞、マクロファージ、線維芽細胞、およびその他を含んでいる。大型黄体細胞は、ペプチド(リラキシン、オキシトシン)を生成し、小細胞よりステロイドよりアロマターゼ活性および大プロゲステロン合成においてより活性です。
プロゲステロンのピークは、LGのピーク後8日目に観察される。黄体期のプロゲステロンとエストラジオールは、LHのパルス出力と相関して散発的に分泌されることが指摘された。黄色体の形成により、インヒビン産生の制御は、FSHからLHに移行する。インギビンは、エストラジオールがLHのピークまで増加すると増加し、LHのピーク後も増加し続け、エストロゲンのレベルは低下する。インヒビンおよびエストラジオールは顆粒膜細胞によって分泌されるが、それらは異なる方法で調節される。黄体期の終わりのインヒビンの減少は、次のサイクルのためのFSHの増加に寄与する。
非常に迅速に黄色の体 - 排卵後9-11日に減少します。
変性のメカニズムは明らかではなく、子宮内膜に見られるように、エストロゲンのルテオリックな役割または受容体結合機構に関連していない。黄色い体によって生成されるエストロゲンの役割についての別の説明がある。子宮内膜におけるプロゲステロン受容体の合成では、エストロゲンが必要であることが知られている。排卵後の子宮内膜におけるプロゲステロン関連の変化にはおそらく黄体期エストロゲンが必要である。不十分なエストロゲン含量の結果として、プロゲステロン受容体の不適切な発達は、おそらく不妊症および初期妊娠喪失の別の機構であり、黄体期の劣性の別の形態である。黄色体の寿命は排卵時に設定されると考えられている。そして妊娠と関連して絨毛性性腺刺激ホルモンが支持されていなければ、それは確実に後退します。したがって、黄色の身体の退行は、エストラジオール、プロゲステロンおよびインヒビンのレベルの低下をもたらす。還元インヒビンは、FSHに対するその阻害効果を除去する。エストラジオールおよびプロゲステロンの減少は、GnRHの分泌を非常に迅速に回復させ、脳下垂体からのフィードバック機構を除去することを可能にする。インヒビンおよびエストラジオールの減少は、GnRHの増加とともに、LHに対してFSHの有病率をもたらす。FSHの増加は、その後の支配的な卵胞の選択とともに卵胞の成長をもたらし、妊娠が起こらない場合には新たなサイクルが始まる。ステロイドホルモンは、生殖生物学および一般的な生理学において主導的役割を果たす。彼らは人の表現型を決定し、心血管系、骨、皮膚の代謝、身体の一般的な健康に影響を与え、妊娠において重要な役割を果たす。ステロイドホルモンの作用は、生理学的応答を誘発するために細胞外シグナルを細胞の核に伝達するために必要な細胞内および遺伝子機構を反映する。
エストロゲンは細胞膜を通って拡散し、細胞の核に位置する受容体に結合する。次いで、受容体 - ステロイド複合体はDNAに結合する。標的細胞において、これらの相互作用は、遺伝子発現、すなわちタンパク質の合成を細胞および組織の特定の機能に導く。