
コレラ菌
最後に見直したもの: 04.07.2025

すべてのiLiveコンテンツは、可能な限り事実上の正確さを保証するために医学的にレビューまたは事実確認されています。
厳格な調達ガイドラインがあり、評判の良いメディアサイト、学術研究機関、そして可能であれば医学的に査読された研究のみにリンクしています。 かっこ内の数字([1]、[2]など)は、これらの研究へのクリック可能なリンクです。
当社のコンテンツのいずれかが不正確、期限切れ、またはその他の疑問があると思われる場合は、それを選択してCtrl + Enterキーを押してください。
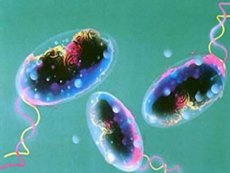
WHOによると、コレラは、コレラ菌(Vibrio cholerae)の感染によって引き起こされる、米のとぎ汁のような便を伴う、急性で重篤な脱水症状を伴う下痢を特徴とする感染症です。流行時に広範囲に蔓延する能力が顕著であること、重篤な経過を辿ること、そして高い死亡率を特徴としていることから、コレラは特に危険な感染症と考えられています。
コレラの歴史的な発祥の地はインド、より正確にはガンジス川とブラマプトラ川のデルタ地帯(現在のインド東部とバングラデシュ)で、太古の昔から存在しています(この地域のコレラの流行は紀元前500年頃から観察されています)。この地域でコレラの流行地が長く存在してきた理由は数多くあります。コレラ菌は水中で長期間生存できるだけでなく、12℃以上の温度や有機物の存在といった好条件下であれば水中で繁殖することができます。これらの条件はすべてインドに備わっています。熱帯気候(年間平均気温25~29℃)、豊富な降雨量と湿地、特にガンジス川デルタ地帯の高い人口密度、水中の多量の有機物、年間を通して続く下水や排泄物による水質汚染、低い物質的な生活水準、住民の独特の宗教儀式や儀礼です。
コレラの流行の歴史には4つの時期が区別できます。
第 1 期 - 1817 年まで、コレラは東アジアと南アジア、主にインドに集中しており、国境を越えて広がることはありませんでした。
第2期 - 1817年から1926年。インドとヨーロッパ諸国、そしてその他の国々との間に広範な経済関係やその他のつながりが確立されたことで、コレラはインドを越えて経済・宗教関係を媒介に蔓延し、6度のパンデミックを引き起こし、数百万人の命を奪いました。ロシアはヨーロッパ諸国の中で最初にコレラが蔓延した国でした。1823年から1926年にかけて、ロシアは57年間にわたりコレラの流行を経験しました。この期間に560万人以上がコレラに罹患し、214万人(全体の40%)が死亡しました。
第三期 - 1926年から1961年まで コレラが再び流行の中心地となり、比較的良好な状態が続きました。飲料水の浄化、廃水の除去と消毒、そして検疫機関の設置を含む特別なコレラ対策のための近代的なシステムの発達により、世界各国は再びコレラの侵攻から確実に守られると思われました。
第四期は1961年に始まり、現在まで続いています。第七期パンデミックはインドではなくインドネシアで始まり、急速にフィリピン、中国、インドシナ諸国、そしてアジア、アフリカ、ヨーロッパの他の国々へと広がりました。このパンデミックの特徴は、第一に、コレラ菌の特殊な変異体であるコレラ菌エルトルによって引き起こされたことです。この菌は1961年までコレラの原因菌として公式に認められていませんでした。第二に、持続期間の点で、それ以前のすべてのパンデミックを凌駕しました。第三に、コレラは2波に分かれて発生し、第1波は1990年まで続き、第2波は1991年に始まり、1866年以来コレラの流行がなかった米国を含む南北アメリカの多くの国を襲った。1961年から1996年までの間に、146か国で3,943,239人がコレラに罹患した。
コレラの原因菌であるコレラ菌は、1883 年の第 5 回パンデミックの際に R. コッホによって発見されましたが、このビブリオ菌は 1854 年に F. パチーニによって下痢患者の排泄物から初めて発見されました。
コレラ菌はビブリオ科に属し、ビブリオ属(ビブリオ、エロモナス、プレシオモナス、フォトバクテリウム)を含む。ビブリオ属には1985年以降25種以上が属しており、その中でヒトにとって最も重要なのはコレラ菌、腸炎ビブリオ、アルギノリティカス菌、ビブリオ・バルニフィカス、ビブリオ・フルビアリス菌である。
ビブリオ属の主な特徴
短い、胞子および莢膜を形成しない、湾曲または直線状のグラム陰性桿菌。直径0.5µm、長さ1.5~3.0µm。運動性(V. cholerae は単毛性で、一部の種は2本以上の極性鞭毛を有する)。通常の培地で良好かつ迅速に増殖し、化学有機栄養体であり、炭水化物を発酵させてガスを発生せずに酸を生成する(グルコースはエムデン・マイヤーホフ経路で発酵される)。酸化酵素陽性で、インドールを生成し、硝酸塩を亜硝酸塩に還元する(V. cholerae はニトロソインドール反応陽性)。ゼラチンを分解し、多くの場合Voges-Proskauer反応陽性(すなわち、アセチルメチルカルビノールを形成)を示し、尿素分解酵素を持たず、H2Sを生成せず、リジンおよびオルニチン脱炭酸酵素を持つが、アルギニンジヒドロラーゼを持たない。ビブリオ属の特徴は、ほとんどの細菌株が薬剤0/129(2,4-ジアミノ-6,7-ジアゾプロピルプテリジン)に感受性を示すことです。一方、シュードモナス科および腸内細菌科の代表的な菌株はこの薬剤に耐性を示します。ビブリオは好気性菌および通性嫌気性菌であり、最適生育温度は18~37℃、pH 8.6~9.0(pH 6.0~9.6の範囲で生育)で、一部の種(好塩菌)はNaClがないと生育しません。DNA中のG + C含有量は40~50モル%(コレラ菌の場合は約47モル%)です。生化学検査は、ビブリオ科内で形態的に類似するエロモナス属およびプレシオモナス属と区別するため、また腸内細菌科と区別するために使用されます。
コレラ菌は、シュードモナス科とは異なり、エムデン・マイヤーホフ経路(酸素の関与なし)のみでグルコースを発酵するのに対し、シュードモナス科は酸素存在下でのみグルコースを消費します。この違いは、ヒュー・ライフソン培地で容易に確認できます。この培地には、寒天、グルコース、指示薬が含まれています。ヒュー・ライフソン培地を用いて2列に播種を行い、そのうちの1列にはワセリン(嫌気性条件を作り出すため)を入れます。コレラ菌が増殖した場合は両方の試験管で培地の色が変わりますが、シュードモナス科が増殖した場合は、ワセリンを入れていない試験管(好気性条件)のみで色が変わります。
コレラビブリオは栄養培地をほとんど必要としません。0.5~1.0%の塩化ナトリウムを含む1%アルカリ性(pH 8.6~9.0)ペプトン水(PV)中では、他の細菌よりも速く、良好に増殖します。プロテウスの増殖を抑制するには、1%PVに亜テルル酸カリウム(最終希釈率1:100,000)を添加することが推奨されます。1%PVはコレラビブリオにとって最適な増菌培地です。増殖後6~8時間でPVの表面に薄く、緩やかな灰色の膜を形成します。この膜は振ると簡単に破壊され、薄片状に底に沈みます。PVは中程度に濁った状態になります。コレラ菌の分離には、アルカリ性寒天、胆汁酸寒天、アルカリ性アルブミン寒天、血液添加アルカリ性寒天、乳糖蔗糖など、様々な選択培地が提案されています。最も優れた培地はTCBS(チオ硫酸クエン酸-ブロモチモール蔗糖寒天)培地とその改良培地です。しかし、最も一般的に使用されるのはアルカリ性MPA培地で、この培地上でコレラ菌は滑らかでガラス状透明、青みがかった色調の円盤状の粘稠なコロニーを形成します。
ビブリオ菌は、ゼラチンの柱に注入して播種すると、22~23℃の温度で2日後に、泡の形で表面から液化を起こし、次に漏斗状の泡になり、最終的には層状に液化します。
牛乳の中では、ビブリオ菌は急速に増殖し、24~48時間後に凝固を引き起こし、その後牛乳のペプトン化が起こり、3~4日後に牛乳のpHが酸性側に変化することでビブリオ菌は死滅します。
B. ハイバーグは、マンノース、スクロース、アラビノースを発酵する能力に基づいて、すべてのビブリオ(コレラ菌とコレラ様菌)をいくつかのグループに分類しました。現在、その数は 8 つに上ります。
コレラ菌はハイベルグの第一グループに属します。
コレラ菌と形態学的、培養学的、生化学的特徴が類似するビブリオは、パラコレラ、コレラ様、NAGビブリオ(非凝集性ビブリオ)、O1群に属さないビブリオなど、様々な名称で呼ばれてきました。O1群に属さないビブリオは、コレラ菌との関連性を最も的確に表しています。A. ガードナーとK. ベンカト=ラマンによって確立されたように、コレラ菌とコレラ様ビブリオは共通のH抗原を持ちますが、O抗原は異なります。O抗原に基づいて、コレラ菌とコレラ様ビブリオは現在139のO血清群に分類されていますが、その数は絶えず増加しています。コレラ菌はO1群に属します。コレラ菌は共通のA抗原と、2つの型特異的抗原(B抗原とC抗原)を有し、これらによって小川血清型(AB型)、稲葉血清型(AC型)、彦島血清型(ABC型)の3つの血清型に区別されます。解離期のコレラ菌はOR抗原を有します。そのため、O血清、OR血清、そして型特異的な稲葉血清と小川血清がコレラ菌の同定に用いられます。
1992年から1993年にかけて、バングラデシュ、インド、中国、マレーシアなどの国々で大規模なコレラ流行が発生しました。その原因菌は、これまで知られていなかったコレラ菌(Vibrio cholerae)の新たな血清型でした。この菌は、O139抗原と多糖体莢膜を有し、他のO血清とは凝集しないという点で、O1型コレラ菌とは抗原特性が異なります。コレラを引き起こす能力、すなわち外毒素であるコレロゲンを合成する能力を含む、その他の形態学的および生物学的特性はすべて、O1型コレラ菌と類似していることが判明しました。その結果、O抗原を変化させた突然変異の結果として、コレラの新たな原因菌であるO139型コレラ菌が出現したと考えられ、ベンガルコレラ菌(V. cholerae O139 bengal)と命名されました。
いわゆるコレラ様ビブリオとコレラ菌(V. cholerae)との関係については、長らく不明瞭な点が残されていました。しかしながら、コレラ菌とコレラ様ビブリオ(NAGビブリオ)を70以上の特徴で比較した結果、両者の類似性は90%に達し、DNA相同性はコレラ菌と研究対象のNAGビブリオの間で70~100%であることが分かりました。したがって、コレラ様ビブリオはコレラ菌と同種とみなされ、主にO抗原が異なることから、非O1群ビブリオ(非O1コレラ菌)と呼ばれています。
コレラ菌は、コレラ菌、エルトール菌、プロテウス菌、アルベンシス菌の4つのバイオタイプに分類されます。エルトール菌の性質については、長年議論されてきました。このビブリオは、1906年にエルトール検疫所のF.ゴットシュリッヒにより、赤痢で死亡した巡礼者の遺体から分離されました。ゴットシュリッヒは、このような菌株をいくつか分離しました。それらは、すべての特性においてコレラ菌と変わらず、コレラのO血清によって凝集しました。しかし、当時、巡礼者の間でコレラは発生しておらず、コレラ菌の長期保有は考えにくいと考えられていたため、コレラにおけるエルトール菌の病因的役割の可能性については、長らく議論の的となっていました。さらに、エルトール菌はコレラ菌とは異なり、溶血作用を持っていました。しかし、1937年にこのビブリオはスラウェシ島(インドネシア)で大規模かつ深刻なコレラの流行を引き起こし、死亡率は60%を超えました。最終的に1961年に第7次パンデミックの原因となり、1962年にそのコレラの性質に関する疑問がようやく解決されました。V. choleraeとV. eltorの違いは、いくつかの特徴に関係するだけです。その他のすべての特性において、V. eltorはV. choleraeと根本的に異なるものではありません。さらに、現在では、V. proteusバイオタイプ(V.finklerpriori)には、以前はNAGビブリオと呼ばれていたO1グループ(現在はO139)を除くビブリオのグループ全体が含まれることが確立されています。V. albensisバイオタイプはエルベ川から分離され、リン光能力を持っていますが、それを失っており、V. proteusと違いはありません。これらのデータに基づき、コレラ菌種は現在、V. cholerae O1 cholerae、V. cholerae eltor、V. cholerae O139 bengal、およびV. cholerae non O1の4つのバイオタイプに分類されています。最初の3つは、2つの血清型O1とO139に属します。最後のバイオタイプには、以前のバイオタイプであるV. proteusとV. albensisが含まれ、O1およびO139血清によって凝集しないビブリオの他の多くの血清型、つまりNAGビブリオによって代表されます。
コレラ菌の病原性因子
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ]
コレラ菌の走化性
これらの特性を利用して、ビブリオ菌は上皮細胞と相互作用します。コレラ菌の変異体(走化性を失ったもの)では毒性が大幅に低下し、Mob変異体(運動性を失ったもの)では毒性が完全に消失するか、大幅に減少します。
ビブリオ菌が微絨毛に付着し、小腸粘膜に定着するための接着因子および定着因子。接着因子には、ムチナーゼ、可溶性ヘマグルチニン/プロテアーゼ、ノイラミニダーゼなどがあり、粘液を構成する物質を破壊することで接着と定着を促進します。可溶性ヘマグルチニン/プロテアーゼは、ビブリオ菌が上皮細胞受容体から分離し、腸管から外部環境へ排出されることを促進し、伝染の拡大を確実にします。ノイラミニダーゼは、コレラ原菌と上皮細胞の結合を強化し、毒素の細胞内への浸透を促進し、下痢の重症度を高めます。
コレラ毒素はコレラ原です。
下痢を引き起こす可能性があるが、コレラゲンとは遺伝的または免疫学的関係がない、いわゆる新しい毒素。
皮膚神経毒性因子および出血性因子。これらの毒性因子の性質とコレラの病因における役割については、十分な研究が行われていない。
[ 13 ], [ 14 ], [ 15 ], [ 16 ], [ 17 ], [ 18 ]
コレラ菌のエンドトキシン
コレラ菌の脂質多糖類は強い内毒素特性を持ち、全身の中毒を引き起こします。
コレラ菌の病原性因子として挙げられている主要なものは、この疾患の病因となる外毒素コレラゲン(CTX AB)です。コレラ分子は、AとBの2つのフラグメントで構成されています。フラグメントAは、A1とA2の2つのペプチドで構成され、コレラ毒素特有の性質を持ち、スーパー抗原としての性質を持っています。フラグメントBは、5つの同一のサブユニットで構成されています。フラグメントBは、1) 腸管上皮細胞の受容体(モノシアロガングリオシド)を認識し、結合する、2) サブユニットAの通過のための膜内疎水性チャネルを形成する、という2つの機能を果たします。ペプチドA2は、フラグメントAとBを結合する役割を果たします。実際の毒性機能は、ペプチドAj(ADPリボシルトランスフェラーゼ)によって実行されます。ペプチドAjはNADと相互作用し、その加水分解を引き起こします。生成されたADPリボースは、アデニル酸シクラーゼの調節サブユニットに結合します。これによりGTP加水分解が阻害されます。結果として生じるGTP + アデニル酸シクラーゼ複合体は、ATP加水分解を引き起こし、cAMPを生成します。(cAMP蓄積のもう一つの経路は、コレラゲンによるcAMPを5-AMPに加水分解する酵素の抑制です。)外毒素合成をコードするctxAB遺伝子の機能発現は、他の多くの病原性遺伝子、特にtcp遺伝子(毒素制御接着線毛(TCAP)の合成をコードする遺伝子)、調節遺伝子toxR、toxS、toxT、hap遺伝子(可溶性ヘマグルチニン/プロテアーゼ)、およびノイラミニダーゼ遺伝子(ノイラミニダーゼ)の機能に依存します。したがって、コレラ菌の病原性の遺伝子制御は複雑です。
結局、コレラ菌の染色体には2つの病原性アイランドが存在する。1つは糸状の中等度の変換ファージCTXφのゲノムであり、もう1つは同じく糸状の中等度の変換ファージVPIcpのゲノムである。これらの病原性アイランドにはそれぞれ、コレラ病原体の病原性を決定する前期で規定される遺伝子のカセットが含まれている。CTXφプロファージは、CTX遺伝子、新規毒素zotおよびaceの遺伝子、ser遺伝子(接着因子合成)、およびortU遺伝子(機能不明の産物の合成)を持っている。このカセットには、nei遺伝子と、プロファージの複製と染色体への組み込みをコードするRS2ファージ領域も含まれている。zot、ace、およびortU遺伝子は、プロファージが病原体の染色体から排除された場合にファージビリオンを形成するために必要である。
VPIcpプロファージは、tcp遺伝子(線毛(TCPAタンパク質)の産生をコード)、toxT、toxR、act遺伝子(追加の定着因子)、移動遺伝子(インテグラーゼおよびトランスポザーゼ)を保有しています。毒性遺伝子の転写は、toxR、toxS、およびtoxTの3つの調節遺伝子によって制御されます。これらの遺伝子は、転写レベルで協調的に、ctxAB、tcp、およびその他の遺伝子を含む20以上の毒性遺伝子の活性を変化させます。主要な調節遺伝子はtoxR遺伝子です。この遺伝子が損傷または欠損すると、非毒性になるか、コレラ毒素CTXおよびTCPAの産生が100分の1以下に減少します。おそらく、これは、温和変換ファージによって形成される病原性島や他の細菌種における毒性遺伝子の協調的な発現を制御する方法であると考えられます。別のプロファージ K139 がコレラ菌の染色体中に存在することは確認されていますが、そのゲノムはほとんど研究されていません。
Hap遺伝子は染色体上に局在しています。したがって、コレラ菌の毒性(病原性)と流行力は、ctxAB、tcp、toxR、hapの4つの遺伝子によって決定されます。
コレラ菌のコレラゲン産生能力を検出するには、さまざまな方法を使用できます。
ウサギを用いた生物学的試験。コレラ菌を乳飲みウサギ(生後2週間以内)に筋肉内注射すると、典型的なコレラ症候群(下痢、脱水症状、そしてウサギの死)が発症します。
PCR、IFM、または受動免疫溶血反応(コレラ原は赤血球のGmjに結合し、抗毒素抗体と補体を加えると溶血する)によるコレラ原の直接検出。しかし、毒素産生能の検出だけでは、そのような株の流行性を判断するには不十分です。そのためには、hap遺伝子の存在を検出する必要があります。したがって、血清群O1およびO139のコレラ菌の毒素産生株と流行性株を区別する最善かつ最も信頼性の高い方法は、ctxAB、tcp、toxR、hapの4つの病原性遺伝子すべてを検出する特異的プライマーを用いたPCRです。
血清群 O1 または O139 以外のコレラ菌がヒトに散発性または群発性下痢性疾患を引き起こす能力は、それぞれアデニル酸シクラーゼまたはグアニル酸シクラーゼ系を刺激する LT 型または ST 型のエンテロトキシンの存在、または ctxAB 遺伝子のみが存在し hap 遺伝子が存在しないことによると考えられます。
第7次パンデミックでは、様々な毒性を持つコレラ菌株が分離されました。コレラ菌株は、胆汁産生性(毒性)、弱胆汁産生性(低毒性)、非胆汁産生性(非毒性)の3種類です。非胆汁産生性コレラ菌は、一般的に溶血活性を示し、コレラ診断ファージHDF(5)によって溶血されず、ヒトに疾患を引き起こしません。
V. cholerae 01(エル・トールを含む)のファージ型別のために、S. Mukherjee はファージセットを提案し、その後ロシアで他のファージが追加されました。このようなファージセット(1~7)を用いることで、V. cholerae 0116 のファージ型を区別することが可能です。毒素産生性および非毒素産生性のV. cholerae エル・トールの識別には、HDF-3、HDF-4、HDF-5 の代わりに、CTX*(毒素産生性エル・トール菌を溶解)およびCTX"(非毒素産生性エル・トール菌を溶解)というファージが現在ロシアで提案されています。
コレラ病原体の耐性
コレラ菌は低温でもよく生き残ります。氷の中では最長 1 か月、海水中では最長 47 日、河川水では 3 ~ 5 日から数週間、沸騰させたミネラルウォーターでは 1 年以上、土壌中では 8 日から 3 か月、新鮮な排泄物では最長 3 日、煮沸した食品 (米、麺類、肉、お粥など) では 2 ~ 5 日、生野菜では 2 ~ 4 日、果物では 1 ~ 2 日、牛乳や乳製品では 5 日間生存します。冷蔵保存すると生存期間が 1 ~ 3 日延長されます。排泄物で汚染されたリネンの上では最長 2 日間、湿った素材の上では 1 週間生存します。コレラ菌は 80 °C の温度では 5 分以内に、100 °C では瞬時に死滅します。酸に非常に敏感です。クロラミンなどの消毒剤の影響下では5~15分以内に死滅します。乾燥や直射日光には弱いものの、有機物に富み、アルカリ性pHで10~12℃以上の水温の開放水域や廃水でも長期間生存し、増殖します。塩素に対する感受性は非常に高く、30分以内に水1リットルあたり0.3~0.4mgの活性塩素を投与することで、コレラ菌を確実に消毒できます。
コレラ菌属に属さない、ヒトに病原性のあるビブリオ
ビブリオ属には25種以上が含まれており、コレラ菌に加えて、腸炎ビブリオ(V. parahaemolyticus)、アルギノリティカス(V. alginolyticus)、バルニフィカス(V. vulnificus)、フルビアリス(V. fluvialis)、フミッシー(V. fumissii)、ミミクス(V. mimicus)、ダムセラ(V. damsela)、ホリサエ(V. hollisae)の少なくとも8種がヒトに疾患を引き起こす可能性があります。これらのビブリオはすべて海や湾に生息し、水泳または魚介類の摂取によって感染します。コレラ菌および非コレラ菌は、胃腸炎だけでなく創傷感染も引き起こすことが判明しています。この能力は、コレラ菌O1群および非O1群、腸炎ビブリオ(V. parahaemolyticus)、アルギノリティカス(V. alginolyticus)、ミミクス(V. mimicus)、ダムセラ(V. damsela)、バルニフィカス(V. vulnificus)で確認されています。海洋動物の殻によって損傷を受けたり、感染した海水と直接接触したりすると、軟部組織に炎症プロセスを引き起こします。
リストされている病原性のないコレラ菌ビブリオの中で、最も実用的な関心が高いのは、V. parahaemolyticus、V. alginolyticus、V. vulnificus、および V. fluvialis です。
腸溶性ビブリオの一種であるV. parahaemolyticusは、1950年に日本で初めて分離されました。このとき、半干しイワシの摂取によって引き起こされた大規模な食中毒が発生し、死亡率は7.5%でした。原因菌は1963年に坂崎龍一によってビブリオ属に分類されました。彼は研究対象の菌株をV. parahaemolyticusとV. alginolyticusの2種に分類しました。両種とも沿岸海水およびその生息域に生息する好塩菌(ギリシャ語でhalsは塩)で、通常のビブリオとは異なり、好塩菌は塩化ナトリウムのない培地では生育せず、高濃度の塩化ナトリウムでよく繁殖します。好塩性ビブリオの種の分類は、スクロースを発酵させ、アセチルメチルカルビノールを形成し、10%塩化ナトリウムのPVで繁殖する能力によって決定されます。これらの特徴はすべて、V. alginolyticus 種に固有のものですが、V. parahaemolyticus には存在しません。
腸炎ビブリオには、熱に不安定な鞭毛H抗原、120℃で2時間加熱しても破壊されない耐熱性O抗原、そして加熱によって破壊される表面K抗原という3種類の抗原があります。腸炎ビブリオの新鮮分離培養物は、明確に定義されたK抗原を有しており、生きたビブリオを相同なO血清による凝集から保護します。H抗原はすべての株で共通していますが、単毛類のH抗原は周毛類のH抗原とは異なります。O抗原に基づいて、腸炎ビブリオは14の血清群に分類されます。血清群内では、ビブリオは K 抗原に応じて血清型に分類され、その総数は 61 です。腸炎ビブリオの抗原スキームは、ヒトから分離された株に対してのみ開発されています。
腸炎ビブリオの病原性は、腸管毒性を持つ溶血素を合成する能力と関連しています。この溶血素は神奈川法を用いて検出されます。その本質は、ヒトに対して病原性のある腸炎ビブリオが、7% NaClを含む血液寒天培地上で明瞭な溶血を引き起こすことにあります。5%未満のNaClを含む血液寒天培地では、腸管毒性を持つ菌株のみが溶血を引き起こします。腸炎ビブリオは、日本海、カスピ海、黒海などの沿岸に生息しています。食中毒や赤痢様疾患を引き起こします。腸炎ビブリオに感染した生または半生の魚介類(海水魚、カキ、甲殻類など)を摂取することで感染します。
上述のコレラ以外の8種のビブリオ菌のうち、ヒトに対して最も病原性が高いのはビブリオ・バルニフィカスです。1976年にベネケア・バルニフィカスとして初めて記載され、1980年にビブリオ・バルニフィカスに再分類されました。海水や海水中に多く存在し、様々なヒト疾患を引き起こします。海洋起源および臨床起源のビブリオ・バルニフィカス株は、表現型的にも遺伝学的にも互いに違いはありません。
V. vulnificus による創傷感染は急速に進行し、腫瘍の形成を招き、続いて組織壊死を引き起こし、発熱、悪寒、時には激しい痛みを伴い、場合によっては切断が必要になります。
V. vulnificus は外毒素を産生することが確認されています。動物実験では、この病原体が浮腫と組織壊死を伴う重度の局所損傷を引き起こし、死に至ることが示されています。この疾患の発症における外毒素の役割は現在研究中です。
V. vulnificus は創傷感染に加え、溺水者に肺炎を引き起こし、海水に曝露した女性に子宮内膜炎を引き起こす可能性があります。V. vulnificus による感染症の中で最も重篤なのは、生牡蠣(および場合によっては他の海産動物)の摂取に関連する一次性敗血症です。この疾患は非常に急速に進行し、患者は倦怠感、発熱、悪寒、衰弱を呈し、その後、重度の低血圧に陥ります。これが主な死因です(死亡率は約50%)。
V. fluvialis は、1981年に初めて胃腸炎の病原体として記載されました。アルギニンジヒドロラーゼは有するが、ネトルニチンおよびリジンデカルボキシラーゼは有さない、コレラ病原性のないビブリオ菌群(V. fluvialis、V. furnissii、V. damsela、すなわち、表現型がAeromonasに類似する)に属します。V. fluvialis は、激しい嘔吐、下痢、腹痛、発熱、および重度または中等度の脱水症状を伴う胃腸炎の一般的な原因菌です。主な病原体はエンテロトキシンです。
コレラの疫学
感染の主な原因は、コレラ患者またはビブリオ菌のキャリアである人間と、それらに汚染された水のみです。自然界では動物がコレラに感染することはありません。感染経路は糞口感染です。感染経路は、a) 主なもの - 飲用水、入浴用水、家庭用水、b) 家庭内接触、c) 食物です。コレラのすべての主要な流行とパンデミックは、水に関連していました。コレラ菌は、人体と開放水域の特定の生態系の両方でその個体群の存在を確保する適応メカニズムを備えています。コレラ菌によって引き起こされる重度の下痢は、腸から競合細菌を排除し、環境、特に廃水とそれらが投棄される開放水域での病原体の広範な拡散に寄与します。コレラに感染した人は、便1mlあたり1億~10億個という膨大な量の病原体を排泄します。ビブリオキャリアは便1mlあたり10万~10万個のビブリオを排泄し、感染量は約100万個です。健康なキャリアにおけるコレラ菌の排泄期間は7~42日、回復した人では7~10日です。これより長い排泄期間は極めてまれです。
コレラの特徴は、発症後、原則として長期の保菌がなく、安定した流行地が形成されないことです。しかし、既に述べたように、大量の有機物、洗剤、食塩を含む廃水による開放水域の汚染により、夏季にはコレラ菌は長期間生存するだけでなく、増殖することさえあります。
疫学的に極めて重要な事実は、O1群コレラ菌(非毒素産生性および毒素産生性)が、培養されていない形態で様々な水生生態系において長期間生存できるという事実です。ポリメラーゼ連鎖反応(PCR)を用いた検査では、CIS諸国における複数の風土病地域の様々な水域において、細菌学的検査で陰性であったにもかかわらず、培養されていないコレラ菌のvct遺伝子が検出されました。
エル・トール・コレラ菌の流行地はインドネシアであり、この第7次パンデミックの原因菌が同国から出現したことは、インドネシアが独立を獲得した後、外の世界との経済的つながりが拡大したことと関係していると考えられている。また、パンデミックの期間と急速な進行、特に第2波は、アジア、アフリカ、アメリカ諸国におけるコレラに対する免疫の欠如とさまざまな社会的混乱に決定的な影響を受けた。
コレラが発生した場合、さまざまな防疫対策が講じられますが、その中でも最優先かつ決定的な対策は、急性および非定型型の患者と健康なビブリオキャリアを積極的に適時に検出し、隔離(入院、治療)することです。感染経路の可能性を防ぐための対策が講じられます。給水(飲料水の塩素処理)には特に注意を払います。食品工場、児童施設、公共の場では衛生状態を遵守します。開放された水域では細菌学的検査を含む厳格な管理を実施し、住民の予防接種を実施します。
コレラの症状
コレラの潜伏期間は数時間から6日間と様々ですが、通常は2~3日です。小腸の内腔に侵入したコレラ菌は、その運動性と粘膜への走化性により、粘液へと向かいます。粘液を通過するために、コレラ菌はノイラミニダーゼ、ムシナーゼ、プロテアーゼ、レシチナーゼといった多くの酵素を産生します。これらの酵素は粘液に含まれる物質を破壊し、コレラ菌が上皮細胞へ移動することを容易にします。コレラ菌は接着によって上皮のグリコカリックスに付着し、運動性を失うと急速に増殖し、小腸の微絨毛に定着します(カラー挿入図101.2を参照)。同時に、大量の外毒素であるコレラゲンを産生します。コレラゲン分子はモノシアロガングリオシドGniに結合します。そして細胞膜を貫通し、アデニル酸シクラーゼ系を活性化します。蓄積したcAMPは腸管上皮細胞から水分、陽イオン、陰イオン(Na、HCO、Kl、Cl)の過剰分泌を引き起こし、コレラ性下痢、脱水、体内の塩分減少を引き起こします。この疾患には3つのタイプがあります。
- 数時間以内に患者の死に至る、激しく重篤な脱水症状を伴う下痢性疾患。
- 症状がそれほど重くない場合、または脱水症状を伴わない下痢の場合。
- 無症候性の病気の経過(ビブリオ菌の保有)。
コレラが重症化すると、患者は下痢を起こし、排便回数が増え、便の量が増えて水っぽくなり、便臭はなくなり、米のだし汁(粘液の残留物と上皮細胞が浮遊する濁った液体)のような外観になります。その後、衰弱させるほどの嘔吐が起こり、最初は腸の内容物が吐き出され、その後、嘔吐物は米のだし汁のような外観になります。患者の体温は平熱以下に低下し、皮膚は青みがかってしわが寄り、冷たくなります。これはコレラ特有の冷淡な症状です。脱水症状の結果、血液が濃くなり、チアノーゼが発生し、酸素欠乏、腎機能の急激な低下、けいれんの発現、意識喪失、そして死亡に至ることがあります。第7次パンデミックにおけるコレラの死亡率は、先進国で1.5%、発展途上国で50%と幅がありました。
感染後の免疫は強力で持続性が高く、再発はまれです。免疫は抗毒素性と抗菌性を持ち、抗体(抗毒素は抗菌抗体よりも長く持続します)、免疫記憶細胞、そして食細胞によって生じます。
コレラの臨床診断
コレラの診断における主要かつ決定的な方法は細菌学的検査です。患者からの検査材料は、便と嘔吐物です。便はビブリオ菌の保有の有無を調べます。コレラで死亡した人からは、小腸と胆嚢の結紮切片を採取して検査します。外部環境からの検査材料としては、開放型貯水池の水や廃水が最も多く使用されます。
細菌学的研究を実施する場合、次の 3 つの条件を満たす必要があります。
- できるだけ早く患者から採取した材料を播種する(コレラ菌は糞便中で短期間生存する)。
- コレラ菌は化学薬品に非常に敏感なので、材料を入れる容器は化学薬品で消毒したり、化学薬品の痕跡を残したりしてはいけません。
- 他者への汚染や感染の可能性を排除します。
培養物は、PVに播種し、同時にアルカリ性MPAまたは任意の選択培地(TCBSが最適)に播種するという手順で分離されます。6時間後、PV上に形成された膜を検査し、必要に応じて2つ目のPVに移します(この場合、コレラ菌の播種率は10%増加します)。PVからアルカリ性MPAに移します。疑わしいコロニー(ガラス状透明)を移して純粋培養物を得ます。純粋培養物は、形態学的、培養学的、生化学的特性、運動性によって同定され、最終的にO型、OR型、稲葉型、小川型の診断凝集血清およびファージ(HDF)を用いて型別されます。迅速診断にはさまざまな選択肢が提案されていますが、最も優れた方法は発光血清学的方法です。本キットは、コレラ菌を試験物質中(または1% PVを含む試験管2本で予備培養後、片方にコレラファージを添加した試験管2本で培養後)に1.5~2時間以内に直接検出することを可能にします。コレラ菌の迅速検出のため、ニジニ・ノヴゴロドIEMは、13種類の生化学検査(オキシダーゼ、インドール、ウレアーゼ、ラクトース、グルコース、スクロース、マンノース、アラビノース、マンニトール、イノシトール、アルギニン、オルニチン、リジン)を含む紙製指示薬ディスクセットを提案しました。これにより、ビブリオ属の菌を、エロモナス属、プレシオモナス属、シュードモナス属、コマモナス属、および腸内細菌科の菌と区別することができます。糞便および環境物質中のコレラ菌の迅速検出には、抗体診断薬を用いたRPGAを使用することができます。環境物体中の非培養コレラ菌を検出するには、ポリメラーゼ連鎖反応法のみが使用されます。
非Ol群コレラ菌が分離された場合は、他の血清群の対応する凝集血清を用いて型別を行う必要があります。下痢症(コレラ様下痢症を含む)の患者から非Ol群コレラ菌を分離する場合は、Ol群コレラ菌の分離の場合と同様の防疫対策が必要です。必要に応じて、PCR法を用いて、これらのコレラ菌における病原性遺伝子ctxAB、tcp、toxR、hapの存在を確認します。
コレラの血清学的診断は補助的な性質を有します。この目的には凝集反応を用いることもできますが、ビブリオシダーゼ抗体または抗毒素の力価を測定する方が適切です(コレラに対する抗体は酵素免疫測定法または免疫蛍光法によって測定されます)。
コレラ以外の病原性ビブリオの臨床診断
非コレラ病原性ビブリオによる疾患の診断の主な方法は、TCBS、マッコンキーなどの選択培地を使用した細菌学的検査です。分離培養物がビブリオ属に属するかどうかは、この属の細菌の主な特徴に基づいて判定されます。
コレラの特異的予防
人工免疫の構築を目的として、不活化コレラワクチン(稲葉株および小川株由来)、皮下注射用コレラトキソイド、そして交差免疫が形成されないため、稲葉株および小川株のアナトキシンと体細胞抗原からなる二価化学ワクチンを含むコレラワクチンが提案されました。しかし、ワクチン接種後の免疫持続期間は6~8か月以内であるため、ワクチン接種は疫学的適応症に基づいてのみ実施されます。コレラ発生地では、抗生物質による予防効果が実証されており、特にコレラ菌が高感受性を示すテトラサイクリンが効果的です。コレラ菌に有効な他の抗生物質も同様の目的で使用できます。