記事の医療専門家

新しい出版物
気管支
最後に見直したもの: 04.07.2025

すべてのiLiveコンテンツは、可能な限り事実上の正確さを保証するために医学的にレビューまたは事実確認されています。
厳格な調達ガイドラインがあり、評判の良いメディアサイト、学術研究機関、そして可能であれば医学的に査読された研究のみにリンクしています。 かっこ内の数字([1]、[2]など)は、これらの研究へのクリック可能なリンクです。
当社のコンテンツのいずれかが不正確、期限切れ、またはその他の疑問があると思われる場合は、それを選択してCtrl + Enterキーを押してください。
右主気管支は気管の延長部であり、長さは28~32mm、内腔径は12~16mmです。左主気管支は長さ40~50mm、幅は10~13mmです。
末梢に向かって、主気管支は葉細気管支、分節細気管支、亜分節細気管支に二分され、さらに末端細気管支と呼吸細気管支へと分岐します。しかし、3つの枝(三分岐)に分岐するなど、それ以上の枝に分岐する場合もあります。
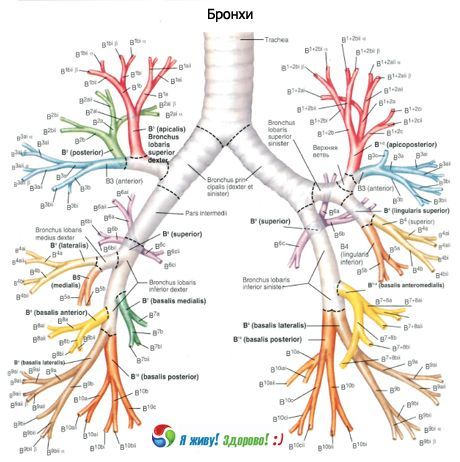
右主気管支は上葉と中間葉に分かれ、中間葉は中葉と下葉に分かれます。左主気管支は上葉と下葉に分かれます。呼吸器系の世代数は様々で、主気管支から肺胞に至るまで、最大で23~26世代に達します。
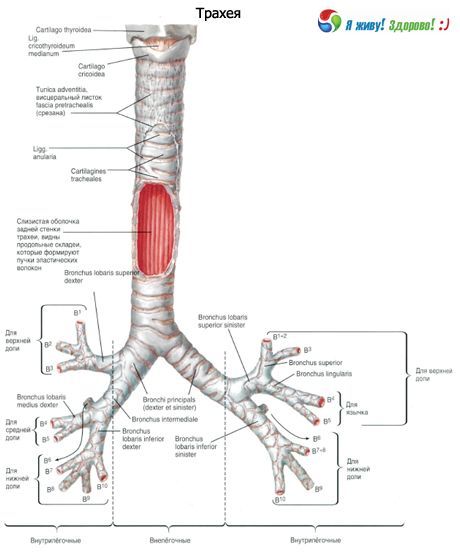
主気管支は第 1 次気管支、小葉気管支は第 2 次気管支、分節気管支は第 3 次気管支などとなります。
第4世代から第13世代までの気管支は直径約2mmで、その数は合計400本です。終末細気管支では、直径は0.5mmから0.6mmの範囲で変化します。喉頭から細気管支までの気道の長さは23cmから38cmです。
右主気管支と左主気管支(気管支原理:右と左)は、第5胸椎の上端の高さにある気管の分岐から始まり、それぞれ右肺と左肺の門まで伸びています。肺門の領域では、各主気管支は小葉(二次)気管支に分岐します。大動脈弓は左主気管支の上にあり、奇静脈は右上にあります。右主気管支はより垂直な位置にあり、左主気管支(長さ4~5cm)よりも短く(約3cm)、右主気管支(直径1.6cm)は左主気管支(1.3cm)よりも広くなっています。主気管支の壁は、気管の壁と同じ構造をしています。主気管支の壁は内側から粘膜で覆われ、外側から外膜で覆われています。壁の基部は軟骨で、後部は閉じられていません。右主気管支には6~8個の軟骨半輪があり、左主気管支には9~12個の軟骨があります。
気管と主気管支の神経支配:左右の反回神経の枝と交感神経幹。
血液供給:下甲状腺動脈、内胸動脈、胸部大動脈の枝。静脈流出は腕頭静脈に行われる。
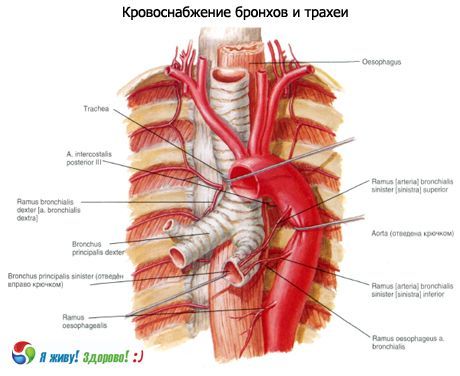
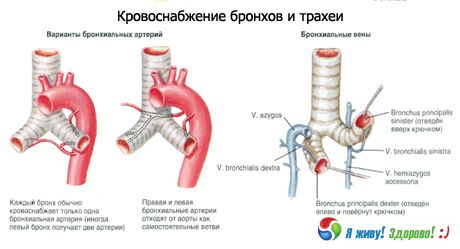
リンパドレナージ:深頸部側方(内頸静脈)リンパ節、気管前および気管傍、気管気管支の上部および下部リンパ節へ。
 [
[ 気管支の組織学的構造
気管と大気管支は、外側から緩い結合組織鞘である外膜で覆われています。外膜は、大気管支内の脂肪細胞を含む緩い結合組織で構成されています。血管、リンパ管、神経が外膜を通過します。外膜は気管支周囲結合組織と明確に区別されておらず、後者と相まって、気管支が肺の周囲の部分に対してある程度変位する可能性があります。
さらに内側には、線維軟骨層と部分的に筋層、粘膜下層、そして粘膜があります。線維層には、軟骨半輪に加えて、弾性繊維のネットワークがあります。気管の線維軟骨膜は、疎性結合組織によって隣接する臓器と結合しています。
気管と大気管支の前壁と側壁は、軟骨とそれらの間に位置する環状靭帯によって形成されています。主気管支の軟骨骨格は、硝子軟骨の半輪で構成されており、気管支の直径が小さくなるにつれてサイズが小さくなり、弾性軟骨の性質を獲得します。したがって、大気管支と中気管支のみが硝子軟骨で構成されています。軟骨は円周の2/3を占め、膜状部分は1/3を占めています。これらは線維軟骨骨格を形成し、気管と気管支の内腔の維持を確保します。
気管と主気管支の膜状部分には、筋束が集中しています。まれな縦線維からなる表層(外層)と、横線維で形成された連続した薄い膜である深層(内層)に区別されます。筋線維は軟骨の両端の間だけでなく、気管の軟骨部分の環状空間にも入り込み、さらに広範囲にわたり主気管支にも入り込んでいます。そのため、気管では、横方向および斜めに配置された平滑筋束は膜状部分にのみ見られ、筋層自体は存在しません。主気管支では、まれな平滑筋群が全周に存在します。
気管支の直径が減少するにつれて、筋層がより発達し、その繊維はわずかに斜めに伸びます。筋肉の収縮は、気管支の内腔を狭めるだけでなく、ある程度短縮も引き起こします。これにより、呼吸器官の容量が減少し、気管支が呼気に参加するようになります。筋肉の収縮により、気管支の内腔は1/4に狭まります。吸入時には、気管支は伸長し、拡張します。筋肉は第2次呼吸細気管支に達します。
筋層の内側には、疎性結合組織からなる粘膜下層があります。粘膜下層には、血管と神経組織、粘膜下リンパ管網、リンパ組織、そして粘液性と漿液性の混合分泌物を伴う管状腺房型の気管支腺の大部分が含まれます。気管支腺は末端部と、粘膜表面にフラスコ状の拡張部を形成する排泄管で構成されています。これらの管が比較的長いことが、腺における炎症過程において気管支炎の長期的経過に寄与しています。腺の萎縮は、粘膜の乾燥や炎症性変化につながる可能性があります。
大きな腺は、気管の分岐部の上方、主気管支が小葉気管支に分岐する領域に最も多く存在します。健康な人は1日に最大100mlの分泌物を分泌します。その95%は水分で、残りの5%はタンパク質、塩分、脂質、無機物で同量です。分泌物の大部分はムチン(高分子糖タンパク質)です。現在、14種類の糖タンパク質が存在し、そのうち8種類が呼吸器系に存在します。
気管支の粘膜
粘膜は、外皮上皮、基底膜、粘膜固有層、粘膜筋板で構成されています。
気管支上皮は高位基底細胞と低位基底細胞から構成され、それぞれが基底膜に付着しています。基底膜の厚さは3.7~10.6μmです。気管と大気管支の上皮は多列で円筒状であり、繊毛を有しています。分節気管支レベルでの上皮の厚さは37~47μmです。上皮は繊毛細胞、杯細胞、中間細胞、基底細胞の4種類の主要な細胞で構成されています。さらに、漿液細胞、刷毛細胞、クララ細胞、クルチツキー細胞も存在します。
繊毛細胞は上皮層の自由表面に多く分布しています(Romanova LK, 1984)。繊毛細胞は不規則な柱状をしており、細胞の中央部には楕円形の泡状の核があります。細胞質の電子光学密度は低く、ミトコンドリアは少なく、小胞体顆粒は未発達です。各細胞は短い微絨毛と、表面に約200本の繊毛を有し、繊毛は厚さ0.3μm、長さ約6μmです。ヒトでは、繊毛の密度は6μm²です。
隣接する細胞の間に空間が形成され、細胞は細胞質とデスモソームの指状の突起によって互いに接続されています。
繊毛細胞の集団は、その頂端表面の分化の程度に応じて、次のグループに分けられます。
- 基底小体と軸糸の形成段階にある細胞。この段階では、繊毛は頂端面に存在しません。この時期に中心小体が集積し、細胞の頂端面へと移動します。そして、繊毛の軸糸が形成される基底小体が形成されます。
- 繊毛形成および繊毛成長が中程度に発現している段階にある細胞。このような細胞の頂端面には少数の繊毛が出現し、その長さは分化細胞の繊毛の長さの1/2~2/3である。この段階では、頂端面では微絨毛が優勢である。
- 繊毛形成と繊毛成長が活発な段階にある細胞。このような細胞の頂端面はほぼ完全に繊毛で覆われており、その大きさは繊毛形成の前段階にある細胞の繊毛の大きさに対応している。
- 繊毛形成と繊毛成長が完了した段階にある細胞。このような細胞の頂端面は、密集した長い繊毛で完全に覆われている。電子回折像では、隣接する細胞の繊毛が同じ方向に向いて湾曲していることが示される。これは粘液繊毛輸送の表現である。
これらの細胞群はすべて、光学電子顕微鏡 (SEM) を使用して撮影した写真ではっきりと確認できます。
繊毛は細胞の頂端にある基底小体に付着しています。繊毛の軸糸は微小管によって形成され、微小管は周縁部に9対(ダブレット)あり、中央には2本の単線(シングレット)があります。ダブレットとシングレットはネキシン線維によって連結されています。ダブレットの片側には2本の短い「ハンドル」があり、その中にはATPエネルギーの放出に関与するATPaseが含まれています。この構造により、繊毛は鼻咽頭方向へ16~17の周波数でリズミカルに振動します。
これらは、上皮を覆う粘膜を毎分約 6 mm の速度で動かし、それによって気管支の継続的な排水機能を確保します。
多くの研究者によれば、繊毛上皮細胞は最終分化段階にあり、有糸分裂によって分裂する能力はない。現代の概念によれば、基底細胞は繊毛細胞へ分化できる中間細胞の前駆細胞である。
杯細胞は繊毛細胞と同様に、上皮層の自由表面まで達する。気管および大気管支の膜様部では、繊毛細胞が最大70~80%を占めるのに対し、杯細胞は20~30%以下を占める。気管および気管支の周囲に軟骨性の半輪状構造が見られる部位では、繊毛細胞と杯細胞の割合が異なる領域が見られる。
- 繊毛細胞が優位である。
- 繊毛細胞と分泌細胞の比率はほぼ等しい。
- 分泌細胞が優位である。
- 繊毛細胞が完全にまたはほぼ完全に存在しない(「非繊毛性」)。
杯細胞は、粘液分泌物を分泌するメロクリン型の単細胞腺です。細胞の形状と核の位置は、分泌の段階と、核上層部が粘液顆粒で満たされる段階によって決まります。粘液顆粒はより大きな顆粒に融合し、低電子密度を特徴とします。杯細胞は細長い形状をしており、分泌物が蓄積すると、基底膜上に基底膜と密接に結合した杯のような形状になります。細胞の広い端は自由表面にドーム状に突出し、微絨毛を備えています。細胞質は電子密度が高く、核は円形で、小胞体は粗面でよく発達しています。
杯細胞は不均一に分布しています。走査型電子顕微鏡による観察では、上皮層の様々な領域に、繊毛上皮細胞のみ、または分泌細胞のみからなる不均一な領域が存在することが明らかになりました。しかし、杯細胞が連続的に集積している領域は比較的少ないです。健常者の分節気管支断面の周囲には、繊毛上皮細胞と杯細胞の比率が4:1~7:1の領域があり、他の領域では1:1となっています。
気管支の遠位部では杯細胞の数が減少する。細気管支では、杯細胞はクララ細胞に置き換えられ、粘液の漿液成分と肺胞下層液の生成に関与する。
小気管支および細気管支では杯細胞は通常存在しませんが、病理学的には現れることがあります。
1986年、チェコの科学者たちは、ウサギの気道上皮に様々な粘液溶解物質を経口投与した場合の反応を研究しました。その結果、粘液溶解物質の標的細胞は杯細胞であることが判明しました。粘液が除去されると、杯細胞は通常変性し、徐々に上皮から剥離します。杯細胞への損傷の程度は投与された物質によって異なり、ラソルバンは最も強い刺激作用を示します。ブロンコリジンおよびブロムヘキシンの投与後、気道上皮において新たな杯細胞の大規模な分化が起こり、杯細胞の過形成が起こります。
基底細胞と中間細胞は上皮層の深部に位置し、自由表面に到達しません。これらは最も分化の遅れた細胞形態であり、そのため主に生理的再生が行われます。中間細胞は細長い形状で、基底細胞は不規則な立方体です。どちらも丸くDNAに富む核と少量の細胞質を有し、基底細胞では細胞質の密度がより高くなります。
基底細胞は繊毛細胞と杯細胞の両方を生み出すことができます。
分泌細胞と繊毛細胞は「粘液繊毛装置」という名前で統合されています。
肺気道における粘液の運動過程は粘液繊毛クリアランスと呼ばれます。粘液繊毛クリアランス(MCC)の機能効率は、繊毛上皮の繊毛運動の頻度と同期性、そして非常に重要な粘液の特性とレオロジー特性、すなわち杯細胞の正常な分泌能力に依存します。
漿液細胞は数が少なく、上皮の自由表面に達し、タンパク質分泌物の小さな電子密度の高い顆粒によって区別されます。細胞質も電子密度が高いです。ミトコンドリアと粗面小胞体はよく発達しています。核は円形で、通常は細胞の中央部に位置します。
分泌細胞、あるいはクララ細胞は、小気管支と細気管支に最も多く存在します。漿液細胞と同様に、小さな電子密度の高い顆粒を含みますが、細胞質の電子密度が低く、滑面小胞体が優勢である点で区別されます。円形の核は細胞の中央部に位置しています。クララ細胞はリン脂質の形成に関与し、サーファクタントの産生にも関与している可能性があります。刺激が増加すると、クララ細胞は杯細胞へと変化すると考えられています。
刷子細胞は自由表面に微絨毛を有するが、繊毛は持たない。細胞質の電子密度は低く、核は楕円形で泡状である。Ham A.とCormack D.(1982)のマニュアルでは、刷子細胞は自ら分泌物を分泌する杯細胞とされている。刷子細胞には、吸収、収縮、分泌、化学受容器など、多くの機能があるとされている。しかしながら、ヒトの気道においてはほとんど研究されていない。
クルチツキー細胞は、気管支樹上皮層基底部に広く分布し、細胞質の電子密度が低いことと、電子顕微鏡および銀浸漬光学顕微鏡で観察される小さな顆粒の存在によって基底細胞と区別されます。クルチツキー細胞は、APUD系の神経分泌細胞に分類されます。
上皮の下には基底膜があり、コラーゲンと非コラーゲン糖タンパク質から構成されています。基底膜は上皮を支え、接着する役割を果たし、代謝と免疫反応に関与しています。基底膜とその下の結合組織の状態が、上皮の構造と機能を決定します。粘膜固有層は、基底膜と筋層の間にある疎性結合組織の層で、線維芽細胞、コラーゲン、弾性繊維を含んでいます。粘膜固有層には血管とリンパ管が含まれています。毛細血管は基底膜に達しますが、貫通することはありません。
気管と気管支の粘膜、特に固有板と腺付近の粘膜下層には、上皮を貫通して内腔へ侵入できる遊離細胞が常に存在します。その中でもリンパ球が優勢で、形質細胞、組織球、肥満細胞(ラブロサイト)、好中球および好酸球は少数です。気管支粘膜におけるリンパ細胞の恒常的な存在は、「気管支関連リンパ組織」(BALT)という特別な用語で呼ばれ、空気とともに呼吸器系に侵入する抗原に対する免疫学的防御反応と考えられています。