記事の医療専門家

新しい出版物
卵巣
最後に見直したもの: 04.07.2025

すべてのiLiveコンテンツは、可能な限り事実上の正確さを保証するために医学的にレビューまたは事実確認されています。
厳格な調達ガイドラインがあり、評判の良いメディアサイト、学術研究機関、そして可能であれば医学的に査読された研究のみにリンクしています。 かっこ内の数字([1]、[2]など)は、これらの研究へのクリック可能なリンクです。
当社のコンテンツのいずれかが不正確、期限切れ、またはその他の疑問があると思われる場合は、それを選択してCtrl + Enterキーを押してください。
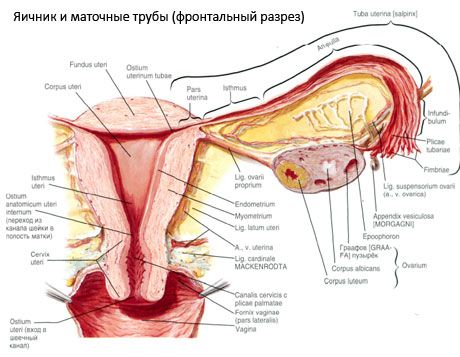
卵巣(ギリシャ語で卵巣)は、子宮広靭帯の後ろの骨盤腔に位置する一対の器官、女性の生殖腺です。卵巣では、女性の生殖細胞(卵子)が発育・成熟し、血液やリンパ液に入る女性ホルモンが生成されます。卵巣は卵形で、前後方向にやや扁平化しています。卵巣の色はピンク色です。出産した女性の卵巣の表面には、排卵と黄体の変化の痕跡である陥没や傷跡が見られます。卵巣の重さは5~8gです。卵巣の大きさは、長さ2.5~5.5cm、幅1.5~3.0cm、厚さ最大2cmです。卵巣には2つの自由表面があります。骨盤腔に面し、部分的に卵管に覆われている内側表面(facies medialis)と、骨盤の側壁に隣接し、わずかに隆起した窪み(卵巣窩)に接する外側表面(facies lateralis)です。この卵巣窩は、上部で腹膜に覆われた外腸骨血管と、下部で子宮動脈と閉鎖動脈の間の角に位置しています。卵巣の背後には、対応する側の尿管が後腹膜を上方から下方にかけて通っています。
卵巣の表面は、凸状の自由縁(後縁)(margo liber)に続き、前方では腸間膜縁(margo mesovaricus)に至り、腹膜(卵巣腸間膜)の短い襞によって子宮広靭帯の後葉に付着しています。この器官の前縁には、溝のある窪み、卵巣門(hilum ovarii)があり、ここから動脈と神経が卵巣に入り、静脈とリンパ管が出ています。卵巣には、卵管に面した丸い上端(extremitas tubaria)と、卵巣靭帯(lig. ovarii proprium)によって子宮につながる下端(extremitas utenna)の2つの端もあります。この靭帯は、約6mmの太さの円形の紐で、卵巣の子宮端から子宮外角まで伸びており、子宮広靭帯の2つの葉の間に位置しています。卵巣靭帯装置には、卵巣吊靭帯(lig.suspensorium ovarii)も含まれます。これは、小骨盤の壁から卵巣まで伸びる腹膜のひだで、内部に卵巣血管と線維性の繊維の束が含まれています。卵巣は短い腸間膜(mesovarium)によって固定されています。腸間膜は、子宮広靭帯の後葉から卵巣の腸間膜縁まで伸びる腹膜の複製です。卵巣自体は腹膜に覆われていません。卵管の最も大きな卵巣縁は、卵巣の管状端に付着しています。卵巣の地形は、子宮の位置と(妊娠中の)子宮の大きさによって決まります。卵巣は、小さな骨盤腔内にある非常に可動性の高い臓器です。
 [
[ 卵巣の血管と神経
卵巣への血液供給は、aa. et vv. ovaricae et uterinae によって行われます。両方の卵巣動脈 (aa. ovaricae dextra et sinistra) は、腎動脈のすぐ下の大動脈の前面から始まり、右側の動脈は通常大動脈から始まり、左側の動脈は腎動脈から始まり、大腰筋の前面に沿って下方および側方に進み、各卵巣動脈は前方で尿管 (尿管に枝を分岐)、外腸骨血管、境界線を横切り、卵巣吊靭帯に位置して骨盤腔に入ります。内側方向に進み、卵巣動脈は卵管の下の子宮広靭帯の葉の間を通り、卵管に枝を分岐させてから卵巣の腸間膜に入ります。それは卵巣の門に入ります。
卵巣動脈の枝は、子宮動脈の卵巣枝と広く吻合しています。卵巣からの静脈流出は、主に卵巣門部に位置する卵巣静脈叢へと行われます。ここから血液は子宮静脈と卵巣静脈の2つの方向に流れ出ます。右卵巣静脈には弁があり、下大静脈に流れ込みます。左卵巣静脈は弁がありませんが、左腎静脈に流れ込みます。
卵巣からのリンパ液の排出はリンパ管を通して行われ、特に卵巣下リンパ叢が明瞭に認められる臓器門付近で多くみられます。その後、リンパ液は卵巣リンパ管に沿って傍大動脈リンパ節へと排出されます。
卵巣の神経支配
交感神経 - 腹腔神経(太陽神経)、上腸間膜神経、下腹神経節後繊維によって供給されます。副交感神経 - 内臓仙骨神経によって供給されます。
卵巣の構造
卵巣の表面は単層の胚上皮で覆われています。その下には、緻密な結合組織タンパク質層(白膜)があります。卵巣の結合組織は、弾性繊維を豊富に含む間質(卵巣間質)を形成します。卵巣の実質である実質は、外層と内層に分かれています。卵巣の中心、門に近い内層は髄質(卵巣髄質)と呼ばれます。この層には、疎な結合組織があり、多数の血管、リンパ管、神経が存在します。卵巣の外層である皮質(卵巣皮質)は、より緻密です。多くの結合組織が含まれており、成熟中の一次卵胞 (卵巣一次卵胞)、二次 (小胞) 卵胞 (卵巣二次卵胞、s.vesiculosi)、成熟卵胞、グラーフ卵胞 (成熟卵胞)、および黄色小体や閉鎖小体が位置しています。
それぞれの卵胞には、女性の生殖卵子、すなわち卵母細胞(オボシトゥス)が含まれています。卵子は直径最大150μmの円形で、核と大量の細胞質を含みます。細胞質には細胞小器官に加え、卵子の栄養源となるタンパク質脂質封入体(卵黄)とグリコーゲンが含まれています。卵子は通常、排卵後12~24時間以内に栄養分を使い果たします。受精が起こらなかった場合、卵子は死滅します。
ヒトの卵子は2つの膜で覆われています。内側には卵子の細胞膜である細胞膜があり、外側には卵子を保護するとともにホルモン形成機能(エストロゲンの分泌)を担う、いわゆる卵胞細胞の層があります。
子宮、卵管、卵巣の生理的位置は、腹膜、靭帯、骨盤組織を繋ぐ懸垂、固定、支持装置によって保たれています。懸垂装置は対をなす構造で、子宮円靭帯、広靭帯、固有靭帯、卵巣懸垂靭帯が含まれます。子宮広靭帯、卵巣固有靭帯、懸垂靭帯は子宮を中央位置に保持します。円靭帯は子宮底を前方に引っ張り、生理的な傾斜を与えます。
固定(アンカー)装置は、子宮を小さな骨盤の中央にしっかりと固定し、左右や前後へのずれを事実上不可能にします。しかし、靭帯装置は子宮下部から伸びているため、子宮が様々な方向に傾く可能性があります。固定装置は、骨盤の遊離組織に位置し、子宮下部から骨盤の側壁、前壁、後壁まで伸びる靭帯、すなわち仙骨靭帯、基底靭帯、子宮膀胱靭帯、膀胱恥骨靭帯で構成されています。
卵巣中膜に加えて、以下の卵巣靭帯が区別されます。
- 卵巣吊靭帯(以前は骨盤漏斗靭帯と呼ばれていた)は、腹膜のひだで、血管(卵巣動脈、卵巣静脈など)、リンパ管、卵巣神経が通っており、骨盤の側壁、腰筋膜(総腸骨動脈の外腸骨動脈と内腸骨動脈の分岐部)、そして卵巣の上端(卵管)の間に張られている。
- 卵巣固有靭帯は、広子宮靭帯の葉の間、後葉に近い位置を通り、卵巣の下端と子宮の側縁を繋いでいます。卵巣固有靭帯は、卵管の始まりと円靭帯の間で、円靭帯の後方および上方で子宮に付着しています。靭帯の厚さには、子宮動脈の終末枝である卵巣動脈が通っています。
- 虫垂卵巣靭帯 この靭帯は、虫垂の腸間膜から右卵巣、または子宮広靭帯まで、腹膜のひだ状に伸びています。この靭帯は不均一で、女性の半分から3分の1に認められます。
支持装置は骨盤底の筋肉と筋膜によって表され、下層、中層、上層(内部)に分かれています。
最も強力なのは上部(内側)の筋層で、肛門を持ち上げる一対の筋肉で代表されます。この層は尾骨から骨盤骨へと3方向(恥骨尾骨筋、腸骨尾骨筋、坐骨尾骨筋)に広がる筋束で構成されています。この筋層は骨盤隔膜とも呼ばれます。
中間層の筋肉は、恥骨結合、恥骨、坐骨の間に位置しています。中間層の筋肉(尿生殖横隔膜)は、尿道と膣が通る骨盤出口の前半を占めています。その中間層の筋肉(尿生殖横隔膜)の間には、外尿道括約筋を形成する筋束があり、後部には横方向に伸びる筋束(会陰深横筋)があります。
骨盤底筋の下部(外側)層は、数字の 8 に似た形状の浅い筋肉で構成されています。これらには、球海綿体筋、坐骨海綿体筋、外肛門括約筋、浅会陰横筋が含まれます。
卵巣の個体発生
卵胞の成長と閉鎖のプロセスは妊娠20週目に始まり、出産時までに最大200万個の卵母細胞が女児の卵巣に残ります。初潮までにその数は30万個に減少します。生殖期間全体を通して、成熟して排卵するのは500個以下の卵胞です。卵胞の初期の成長はFSH刺激に依存せず、限られており、すぐに閉鎖が発生します。ステロイドホルモンの代わりに、局所オートクリン/パラクリンペプチドが一次卵胞の成長と閉鎖の主な調節因子であると考えられています。卵胞の成長と閉鎖のプロセスは、いかなる生理学的プロセスによっても中断されないと考えられています。このプロセスは、子宮内期や閉経期を含むすべての年齢で継続し、妊娠、排卵、無排卵によって中断されます。卵胞の成長を誘発するメカニズムと、各特定の周期における卵胞の数はまだ不明です。
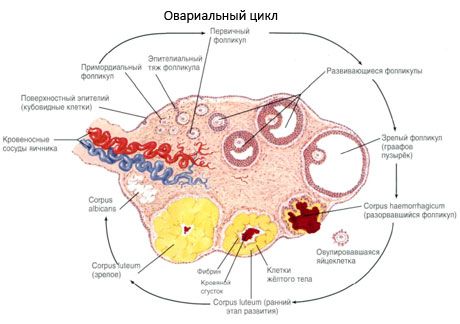
卵胞は発達の過程で複数の段階を経ます。始原生殖細胞は卵黄嚢(尿膜)の内胚葉から発生し、妊娠5~6週で胚の生殖器領域へと移動します。妊娠6~8週から16~20週にかけて続く急速な有糸分裂の結果、胚の卵巣では、薄い顆粒膜細胞層に囲まれた最大600万~700万個の卵母細胞が形成されます。
前胞状卵胞 - 卵母細胞は膜(透明帯)に囲まれています。卵母細胞を取り囲む顆粒膜細胞は増殖を開始し、その成長はゴナドトロピンに依存し、エストロゲンレベルと相関します。顆粒膜細胞はFSHの標的です。前胞状卵胞期の顆粒膜細胞は、主にアンドロゲンをエストラジオールに変換する主要酵素であるアロマターゼの活性を誘導する3種類のステロイドを合成することができます。エストラジオールは自身の受容体の数を増加させ、FSHとは独立して顆粒膜細胞に直接的な分裂促進効果を発揮すると考えられています。エストラジオールは、芳香族化プロセスの活性化を含むFSHの効果を高めるパラクリン因子と考えられています。
FSH受容体は、卵胞の成長が始まるとすぐに顆粒膜細胞の膜上に出現します。FSHの減少または増加は、その受容体の数の変化につながります。このFSHの作用は成長因子によって調節されます。FSHはGタンパク質、アデニル酸シクラーゼ系を介して作用します。卵胞におけるステロイド生成は主にFSHによって制御されますが、このプロセスにはイオンチャネル、チロシンキナーゼ受容体、セカンドメッセンジャーのホスホリパーゼ系など、多くの因子が関与しています。
初期卵胞発育におけるアンドロゲンの役割は複雑です。顆粒膜細胞はアンドロゲン受容体を有しています。これらは、FSH誘導性のエストロゲンへの芳香化の基質となるだけでなく、低濃度でも芳香化を促進することができます。アンドロゲン濃度が上昇すると、前胞状顆粒膜細胞はエストロゲンへの芳香化経路ではなく、5α還元酵素を介したより単純なアンドロゲン経路を優先的に選択し、エストロゲンに変換できないアンドロゲンに変換することで、アロマターゼの活性を阻害します。このプロセスはFSHおよびLH受容体の形成も阻害し、卵胞発育を阻害します。
芳香化過程において、アンドロゲンレベルの高い卵胞は閉鎖過程を経ます。卵胞の成長と発達は、アンドロゲンをエストロゲンに変換する能力に依存します。
FSHが存在する場合、卵胞液の主な物質はエストロゲンです。FSHが存在しない場合は、アンドロゲンです。LHは通常、周期の半ばまで卵胞液中に存在しません。LHレベルが上昇するとすぐに、顆粒膜細胞の有糸分裂活動が低下し、退化性変化が現れ、卵胞内のアンドロゲンレベルが上昇します。卵胞液中のステロイドレベルは血漿中よりも高く、卵巣細胞(顆粒膜細胞と莢膜細胞)の機能活動を反映しています。FSHの唯一の標的が顆粒膜細胞である場合、LHは莢膜細胞、間質細胞と黄体細胞、顆粒膜細胞など、多くの標的を持っています。顆粒膜細胞と莢膜細胞はどちらもステロイド生成能力がありますが、アロマターゼ活性は顆粒膜細胞で優勢です。
LH に反応して卵胞膜細胞はアンドロゲンを生成し、これが顆粒膜細胞によって FSH 誘導による芳香化を通じてエストロゲンに変換されます。
卵胞が成長するにつれて、卵胞膜細胞はLH受容体、P450sec、3β-ヒドロキシステロイドデヒドロゲナーゼの遺伝子を発現し始め、インスリン様成長因子(IGF-1)はLHと相乗的に遺伝子発現を増加させますが、ステロイド生成を刺激しません。
卵巣のステロイド生成は常にLHに依存します。卵胞が成長するにつれて、莢膜細胞はコレステロールからアンドロゲンを生成するP450c17酵素を発現します。顆粒膜細胞はこの酵素を持たず、アンドロゲンからエストロゲンを生成するために莢膜細胞に依存しています。ステロイド生成とは異なり、卵胞形成はFSHに依存します。卵胞が成長しエストロゲンレベルが上昇すると、フィードバック機構が活性化されます。FSH産生が抑制され、卵胞のアロマターゼ活性が低下し、最終的にはアポトーシス(プログラム細胞死)によって卵胞閉鎖に至ります。
エストロゲンとFSHのフィードバック機構は、成長を開始した卵胞の発育を阻害しますが、主席卵胞の発育は阻害しません。主席卵胞にはより多くのFSH受容体が存在し、顆粒膜細胞の増殖とアンドロゲンからエストロゲンへの芳香族化を促進します。さらに、パラクリン経路とオートクリン経路は、胞状卵胞の発育を調節する重要な役割を担っています。
オートクリン/パラクリン制御因子は、顆粒膜細胞によってFSHに反応して合成され、卵胞液中に排出されるペプチド(インヒビン、アクチビン、フォリスタチン)から構成されています。インヒビンはFSH分泌を抑制し、アクチビンは下垂体からのFSH放出を刺激し、卵巣におけるFSHの作用を増強します。フォリスタチンは、おそらくアクチビンに結合することでFSHの活性を抑制します。排卵および黄体の発達後、インヒビンはLHの制御下にあります。
卵巣細胞の成長と分化は、インスリン様成長因子(IGE)の影響を受けます。IGF-1は顆粒膜細胞に作用し、環状アデノシン一リン酸(cAMP)、プロゲステロン、オキシトシン、プロテオグリカン、インヒビンの増加を引き起こします。
IGF-1は卵胞膜細胞に作用し、アンドロゲン産生を増加させます。卵胞膜細胞は、同様にFSHによって調節される腫瘍壊死因子(TNF)と上皮成長因子(EGF)を産生します。
EGFは顆粒膜細胞の増殖を刺激します。卵胞液中の主な成長因子はIGF-2で、IGF-1、TNF-α、TNF-3、EGFも含まれています。
卵巣機能の傍分泌および/または自己分泌調節の破壊は、排卵障害および多嚢胞性卵巣の発達に影響を与えると思われます。
胞状卵胞が成長するにつれて、卵胞液中のエストロゲン含有量が増加します。エストロゲン含有量の増加がピークに達すると、顆粒膜細胞にLH受容体が出現し、顆粒膜細胞の黄体化が起こり、プロゲステロン産生が増加します。このように、排卵前期には、エストロゲン産生の増加がLH受容体の出現を促し、LHが顆粒膜細胞の黄体化とプロゲステロン産生を引き起こします。プロゲステロンの増加はエストロゲンレベルを低下させ、これが月経周期の半ばにFSHの第二ピークを引き起こすと考えられます。
排卵は、LHピークの10~12時間後、およびエストラジオールピークの24~36時間後に起こると考えられています。LHは、卵母細胞における還元分裂、顆粒膜細胞の黄体化、そして卵胞におけるプロゲステロンとプロスタグランジンの合成を刺激すると考えられています。
プロゲステロンはタンパク質分解酵素の活性を高め、プロスタグランジンと共に卵胞壁の破裂に関与します。プロゲステロンによって誘導されるFSHのピークは、プラスミノーゲンをタンパク質分解酵素プラスミンに変換することで卵母細胞が卵胞から脱出することを可能にし、黄体期の正常な発育に必要な数のLH受容体を供給します。
排卵後3日以内に顆粒膜細胞が大きくなり、その中に色素であるルテインで満たされた特徴的な空胞が現れます。黄体莢膜細胞は黄体莢膜と間質から分化し、黄体の一部になります。黄体を貫通する毛細血管は血管新生因子の影響下で急速に発達し、血管新生が改善されると、プロゲステロンとエストロゲンの産生が増加します。ステロイド生成の活性と黄体の寿命は、LHのレベルによって決まります。黄体は均質な細胞構造ではありません。2種類の黄体細胞に加えて、内皮細胞、マクロファージ、線維芽細胞などが含まれています。大きな黄体細胞はペプチド(リラキシン、オキシトシン)を産生し、小さな細胞よりもステロイド生成が活発で、アロマターゼ活性とプロゲステロン合成がより多く見られます。
プロゲステロンのピークは、LHピークから8日目に発生します。黄体期には、LHの脈動的な分泌量と相関して、プロゲステロンとエストラジオールが断続的に分泌されることが知られています。黄体形成に伴い、インヒビン産生の制御はFSHからLHに移行します。インヒビンは、LHピーク前のエストラジオールの上昇に伴って増加し、LHピーク後もエストロゲンレベルが低下するにもかかわらず増加し続けます。インヒビンとエストラジオールは顆粒膜細胞から分泌されますが、それぞれ異なる経路によって制御されています。黄体期末のインヒビンの減少は、次の周期のFSHの上昇に寄与します。
黄体は排卵後9~11日目に急速に減少します。
変性のメカニズムは不明であり、エストロゲンの黄体崩壊作用や子宮内膜で観察されるような受容体関連のメカニズムとは関係がありません。黄体で産生されるエストロゲンの役割については別の説明があります。子宮内膜のプロゲステロン受容体の合成にはエストロゲンが必要であることが知られています。黄体期のエストロゲンは、排卵後の子宮内膜のプロゲステロン関連の変化におそらく必要です。エストロゲンレベル不足の結果としてのプロゲステロン受容体の不十分な発達は、不妊症や早期流産、つまり黄体機能不全の別の形に対するさらなるメカニズムである可能性があります。黄体の寿命は排卵時に決定されると考えられています。そして、妊娠によるヒト絨毛性ゴナドトロピンによるサポートがなければ、黄体は確実に退縮します。したがって、黄体の退行は、エストラジオール、プロゲステロン、インヒビンのレベルの低下をもたらします。インヒビンの減少は、FSHに対するその抑制効果を排除します。エストラジオールとプロゲステロンの減少は、GnRH分泌の急速な回復と下垂体からのフィードバック機構の除去を可能にします。インヒビンとエストラジオールの減少は、GnRHの増加と相まって、LHよりもFSHを増加させます。FSHの増加は卵胞の成長を促し、続いて優位卵胞が選択され、妊娠が起こらない場合は新しい周期が始まります。ステロイドホルモンは、生殖生物学と一般生理学において主導的な役割を果たしています。ステロイドホルモンは、ヒトの表現型を決定し、心血管系、骨代謝、皮膚、一般的な健康状態に影響を与え、妊娠において重要な役割を果たします。ステロイドホルモンの作用は、細胞外シグナルを細胞核に伝達して生理学的反応を引き起こすために必要な細胞内および遺伝子のメカニズムを反映しています。
エストロゲンは細胞膜を拡散透過し、細胞核にある受容体に結合します。その後、受容体-ステロイド複合体はDNAに結合します。標的細胞において、これらの相互作用は遺伝子発現、タンパク質合成、そして特定の細胞および組織の機能につながります。