記事の医療専門家

新しい出版物
クラミジア
最後に見直したもの: 06.07.2025

すべてのiLiveコンテンツは、可能な限り事実上の正確さを保証するために医学的にレビューまたは事実確認されています。
厳格な調達ガイドラインがあり、評判の良いメディアサイト、学術研究機関、そして可能であれば医学的に査読された研究のみにリンクしています。 かっこ内の数字([1]、[2]など)は、これらの研究へのクリック可能なリンクです。
当社のコンテンツのいずれかが不正確、期限切れ、またはその他の疑問があると思われる場合は、それを選択してCtrl + Enterキーを押してください。
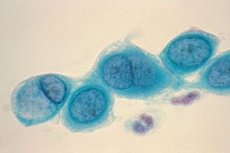
クラミジアは、クラミジア目クラミジア科に属する、グラム陰性の小型球状寄生細菌です。現在、この科には、抗原構造、細胞内封入体、およびスルホンアミド系薬剤への感受性が異なる2つの属が含まれます。クラミジア属(クラミジア・トラコマティス)とクラミドフィラ属(クラミジア・ニューモニア、クラミジア・シッタシ)です。
「クラミジア」(ギリシャ語のchtamys(マントル)に由来)という名前は、微生物粒子の周囲に膜が存在することを反映しています。
すべての種類のクラミジアは共通の形態学的特徴、共通の群抗原、そして異なる繁殖周期を有します。クラミジアはATP合成能力を失ったグラム陰性細菌と考えられており、したがって、絶対細胞内エネルギー寄生虫です。
クラミジア・トラコマティスとクラミジア・ニューモニエは、ヒトに対して明確な病原性を持つ微生物に分類され、人為的クラミジアの原因物質となります。病原体の種類と感染経路(呼吸器系、泌尿生殖器系)によって、呼吸器クラミジアと泌尿生殖器クラミジアに区別されます。
クラミジア・トラコマティスによって引き起こされる病態学的病型は20種類以上報告されており、トラコーマ、結膜炎、鼠径リンパ肉芽腫症、ライター症候群、泌尿生殖器クラミジアなどが含まれます。WHOの推定によると、クラミジア・トラコマティスによる感染症は、性感染症の中でトリコモナス感染症に次いで2番目に多いとされています。世界中で毎年約5,000万人が登録されています。
クラミドフィラ肺炎は、上気道疾患である重度の肺炎を引き起こします。クラミドフィラ肺炎が動脈硬化症や気管支喘息の発症に関与している可能性が示唆されています。
クラミドフィラ・シッタシは、人獣共通感染症であるオウム病の原因です。
クラミジアの形態学的および染色学的特性
クラミジアは球形または卵形の小さなグラム陰性細菌です。鞭毛や莢膜を持ちません。クラミジアの検出には主にロマノフスキー・ギムザ染色法が用いられます。染色の色はライフサイクルの段階によって異なります。基本環は青色の細胞質を背景に紫色に染まり、網状小体は青色に染まります。
細胞壁の構造はグラム陰性細菌に類似していますが、いくつかの違いがあります。典型的なペプチドグリカンは含まれておらず、N-アセチルムラミン酸は細胞壁の構成成分として全く存在しません。細胞壁は外膜で構成されており、LPSとタンパク質が含まれています。ペプチドグリカンが存在しないにもかかわらず、クラミジアの細胞壁は硬く、細胞質は内膜によって制限されています。
クラミジアの外膜(OM)の解析により、外膜の主要タンパク質であるLPS(MOMP)と、OMの内表面に会合するシステインに富むタンパク質OmplおよびOmp3が含まれていることが明らかになりました。クラミジア・プシッタシおよびクラミジア・トラコマティスのLPSとMOMPは、クラミジア・ニューモニエのMOMPとは対照的に、細胞の外表面に局在しています。クラミジア・プシッタシおよびクラミジア・ニューモニエの分子量90~100 kDaのOmpタンパク質も、ここに局在しています。
クラミジアは多形性を有し、これはその繁殖の特殊性に関連しています。クラミジアの独特な(二相)発育サイクルは、感染型(基本小体 - EB)と栄養型(網状小体、または始原小体 - RT)という2つの異なる存在形態が交互に現れることを特徴としています。
微生物にはRNAとDNAが含まれています。RTではRNAがDNAの4倍含まれています。これらの含有量は同等です。
網状体は楕円形、三日月形、双極性桿体、球桿体の形をとり、大きさは300~1000nmです。網状体は感染性はなく、分裂することでクラミジアの増殖を促進します。
楕円形を呈し、大きさは250~500nmの素小体は感染性を有し、感受性細胞に侵入して発育サイクルが進行します。また、緻密な外膜を有するため、細胞外環境において耐性を示します。
クラミジアの培養
クラミジアは絶対寄生虫であるため、人工栄養培地では増殖できず、生細胞内でのみ培養できます。クラミジアはエネルギー寄生虫であり、自力でエネルギーを蓄積して宿主細胞のATPを利用することができません。クラミジアは、HeLa細胞、マッコイ細胞、鶏胚の卵黄嚢、そして感受性動物の体内で35℃の温度で培養されます。
 [ 3 ], [ 4 ], [ 5 ], [ 6 ], [ 7 ], [ 8 ], [ 9 ]
[ 3 ], [ 4 ], [ 5 ], [ 6 ], [ 7 ], [ 8 ], [ 9 ]
クラミジアの抗原構造
クラミジアには、特異抗原(すべてのタイプのクラミジアに共通)- LPS、情報特異抗原(すべてのタイプのクラミジアで異なる)- 外膜に位置する天然のタンパク質、タイプ特異(クラミジア・トラコマティスの血清型で異なる)- 微生物の細胞壁で増殖する LPS、タンパク質の性質を持つ変異体特異抗原の 3 種類の抗原があります。
A、B、C血清型はトラコーマを引き起こすため、眼性クラミジアと呼ばれます。D、E、K、O、H、I、J、K(性器性)血清型は、泌尿生殖器クラミジアとその合併症の原因菌です。L血清型は性器リンパ肉芽腫症の原因菌です。呼吸器クラミジアの原因菌であるクラミジア・ニューモニエには、TWAR、AR、RF、CWLの4つの血清型があります。クラミジア・プシッタシには13の血清型があります。
[ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ], [ 16 ], [ 17 ], [ 18 ]
クラミジアの細胞向性
クラミジア・トラコマティスは尿路生殖管粘膜に向性があり、粘膜上に局所的に留まることも、組織全体に広がることもあります。性病性リンパ肉芽腫の原因菌はリンパ組織に向性があります。
クラミジア・ニューモニエは肺胞マクロファージ、単球、血管内皮細胞で複製され、感染が全身に広がる可能性もあります。
クラミジア・シッタシは単核食細胞を含むさまざまな種類の細胞に感染を引き起こします。
クラミジアのライフサイクル
クラミジアの発育サイクルは 40 ~ 72 時間続き、形態的および生物学的特性が異なる 2 つの存在形態が含まれます。
感染過程の第一段階では、感受性宿主細胞の細胞膜にクラミジア素小体が静電気力の作用によって吸着されます。クラミジアはエンドサイトーシスによって細胞内に導入されます。細胞膜に吸着された素小体は、貪食小胞の形成を伴い細胞質内に陥入します。この段階は7~10時間続きます。
その後、6~8時間以内に、感染性の基本小体は、代謝活性を持ち、非感染性の栄養型細胞内小体(RT)へと再編成され、分裂を繰り返します。これらの細胞内小体は微小コロニーであり、クラミジア封入体と呼ばれます。18~24時間以内に、封入体は宿主細胞膜から形成された細胞質小胞内に局在します。封入体には、100~500個のクラミジア網状小体が含まれることがあります。
次の段階である36~42時間では、成熟(中間体の形成)と網状体の分裂による基本小体への変化が起こります。感染細胞は破壊され、基本小体は細胞から排出されます。細胞外に存在する基本小体は、40~72時間後に新たな宿主細胞に侵入し、クラミジアの新たな発育サイクルが始まります。
このような繁殖サイクルに加えて、不利な条件下では、クラミジアと宿主細胞の間に相互作用を起こす他のメカニズムも発現します。具体的には、ファゴソームにおけるクラミジアの破壊、L型形質転換、そして持続感染です。
クラミジアの変形した形態および持続形態は、元の(網状の)形態に戻り、その後、基本小体へと変形する可能性があります。
宿主細胞の外では代謝機能は最小限に抑えられます。
クラミジアの病原性因子
クラミジアの付着性は、細胞の外膜タンパク質によるもので、このタンパク質は抗貪食作用も有しています。さらに、微生物細胞はエンドトキシンを保有し、エキソトキシンを産生します。エンドトキシンの代表例としてLPSが挙げられますが、これはグラム陰性細菌のLPSと多くの点で類似しています。この物質の熱不安定性はエキソトキシンによって構成されており、あらゆる細菌に存在し、静脈内投与するとマウスに死をもたらします。
クラミジアにはタイプ III 分泌システムがあることがわかっており、このシステムを通じてクラミジアタンパク質が感染プロセスの不可欠な部分として宿主細胞の細胞質に注入されます。
熱ショックタンパク質(HSP)には自己免疫反応を誘発する能力があります。
[ 29 ], [ 30 ], [ 31 ], [ 32 ], [ 33 ], [ 34 ]
クラミジアの生態と耐性
クラミジアは非常に一般的な微生物です。200種以上の動物、魚類、両生類、軟体動物、節足動物から発見されています。形態的に類似した微生物は高等植物からも発見されています。クラミジアの主な宿主はヒト、鳥類、哺乳類です。
クラミジアの原因物質は外部環境下では不安定で、高温に非常に敏感であり、乾燥すると急速に死滅します。50℃では30分後、90℃では1分後に不活化されます。室温(18~20℃)では、病原体の感染力は5~7日後に低下します。37℃では、恒温槽で6時間放置すると毒性が80%低下することが観察されます。低温(-20℃)は、病原体の感染特性の長期保存に寄与します。クラミジアは、紫外線、エチルエーテルおよび70%エタノールとの接触、2%リゾール10分間、2%クロラミンの影響下で急速に死滅します。